
Concepts of Hormone Secretion
Hormones are molecules produced by the endocrine system that send messages to various parts of the body. They help to regulate bodily functions including metabolism, reproduction, growth, and mood. You may be asking yourself: how do hormones elicit a response? The endocrine system uses four different types of hormone signaling to transmit information throughout the body: autocrine, paracrine, endocrine, and neurocrine signaling. This section will examine all four of these mechanisms in addition to discussing the general classes and characteristics of hormones. Lastly, this section will examine how hormone secretion is regulated within our body to prevent hormonal imbalances.
Learning Outcomes
In this section you will learn…
- To differentiate between the two main forms of communication within the body: hormonal and neural.
- The different types of hormone signaling within the endocrine system.
- How different classes of hormones are stored and interact with their corresponding target receptors.
- The general characteristics of all hormones.
- The different form of control over hormone regulation within the human body.
Overview of Communication Systems
The human body has two main systems of communication: the nervous system and the endocrine system. These two systems work together to maintain homeostasis and allow for the coordination of bodily functions.
The Nervous System
As discussed previously in the Neuromuscular Communications unit, the nervous system is the faster out of the two communication methods, transmitting information at rapid speeds from milliseconds to seconds. The nervous system is able to transmit information quickly within the body using neurons to conduct electrical signals and secrete neurotransmitters, such as norepinephrine, to communicate with other cells. Secreted chemical neurotransmitters can then bind to receptors in postsynaptic cells, evoking signaling cascades, and in turn, cellular responses. The signaling between a neuron and its postsynaptic cells continues until the chemical neurotransmitter is no longer secreted from the neuron, ceasing the response from the other cells. This type of rapid communication allows the body to perform quick movements and respond to sudden disruptions in the internal or external environment.[1]
The Endocrine System
The endocrine system consists of cells, tissues, and organs that secrete hormones as either a primary or secondary function. Specifically, the endocrine system includes the pituitary, thyroid, parathyroid, adrenal, and pineal glands. The human body does not always require rapid responses to disruptions in the internal or external environment. Therefore, the body utilizes the endocrine system to regulate disruptions that benefit from a more gradual and elongated response from seconds to minutes. Similar to how the nervous system uses chemicals called neurotransmitters to communicate within the body, the endocrine system uses hormones. The endocrine system releases very low concentrations of hormones from secretory cells into the extracellular space. The hormones diffuse from the extracellular space into the bloodstream where they are transported to all parts of the body to interact with specific ligand receptors. These specific ligand receptors will either be located in the plasma membrane, cytosol or nucleus depending on the nature of the hormone. Receptors for non-lipid soluble (NLS) hormones are located at the plasma membrane because they are unable to cross the phospholipid bilayer of the cell membrane. Meanwhile, since lipid-soluble (LS) hormones can easily cross the phospholipid bilayer, their respective ligand receptors are located either in the cytosol or in the nucleus. Below is a visual representation of NLS and LS hormones travel and bind to their specific ligand receptors to elicit a cellular response. Take note that this learning object refers to NLS hormones as water-soluble hormones. Additionally, although not displayed, LS hormones can also bind to specific ligand receptors on the nuclear membrane.
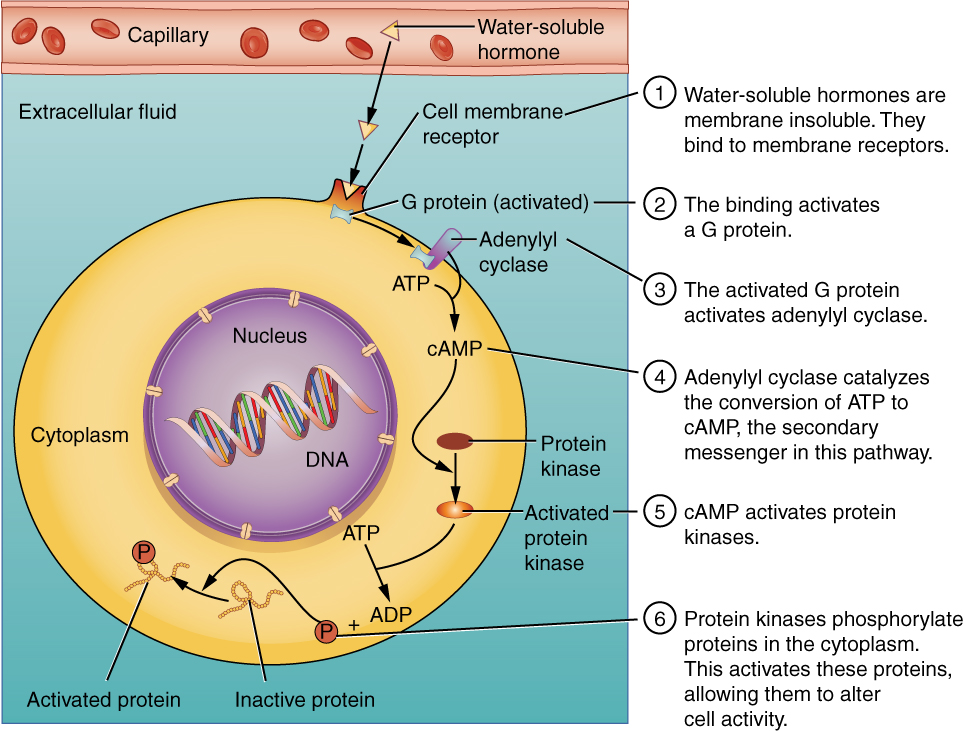
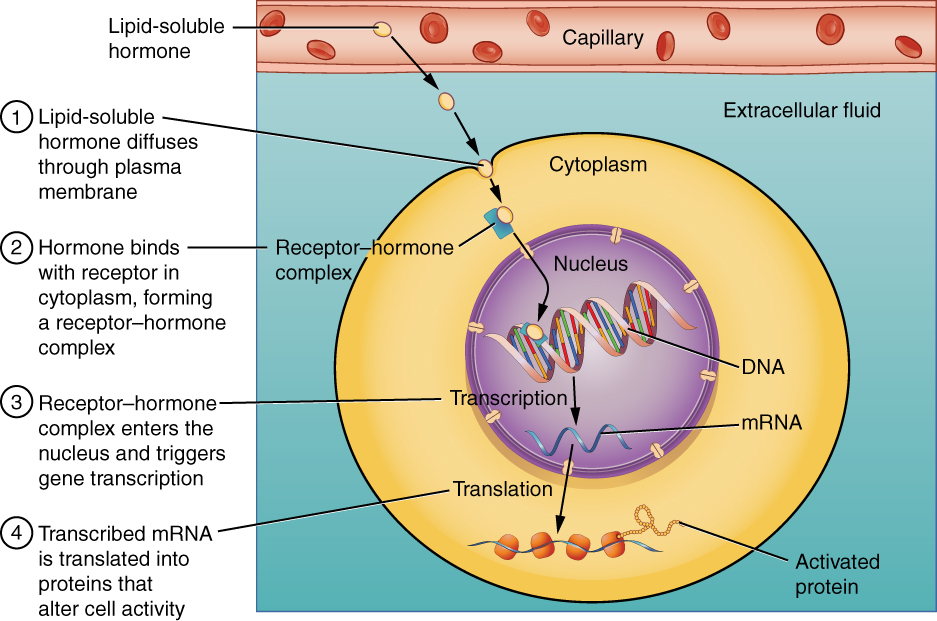
Tips From Past Students
To help you remember where specific hormone ligand receptors will be located think of the phospholipid bilayer of the cell as oil. If a NLS hormone, or in other words a water-soluble hormone, comes into contact with the phospholipid bilayer they will naturally repel on another much like oil and water. As a result, since NLS hormones are unable to cross the phospholipid bilayer they require receptors located on the plasma membrane in order to elicit a response within the cell.
Comparison Between Communication Systems
As mentioned previously, the nervous system communicates via neurotransmitters while the endocrine system utilizes hormones. It is important to note that some specific signaling chemicals (e.g. epinephrine) can act as both a neurotransmitter and a hormone. Epinephrine is classified as a neurotransmitter when it is released by the nervous system to help neurons communicate with one another. However, epinephrine can also be classified as a hormone when it is released by the adrenal glands and has functions peripherally. Therefore, even though some signaling chemicals are used by both systems, hormone-induced responses last longer and require more time to take effect compared to neuronal responses. To better differentiate between the two forms of communication within the body, below is a chart that compares the similarities and differences between the endocrine and nervous systems.
| Communication Medium | Endocrine System | Central Nervous System |
|---|---|---|
| Signalling Chemical | Hormones | Neurotransmitters |
| Chemical Receptor Location(s) | Plasma membrane, cytosol, or nucleus | Postsynaptic neuron membrane |
| Response Times | Seconds to minutes | Milliseconds to seconds |
| Response Control | Involuntary | Voluntary and involuntary |
| Response Location | Local and distant | Local |
To help solidify our understanding of the endocrine system watch the following video. Many of the key points regarding hormonal communication previously discussed are summarized within the first 2 minutes. However, consider watching the full video since it may be helpful to familiarize yourself with the other concepts discussed before continuing to read the rest of the material. Keep in mind that the video is quite simple and does not cover all course concepts!
“The Endocrine System” by Project NEURON Creative, Commons Attribution License (reuse allowed).
Test Your Knowledge
Real-Life Scenario:
Testosterone is a lipid-soluble hormone with a variety of functions in the male body. One of its roles is maintaining bone mass. Consider if and how testosterone would be affected if the quantity of its associated carrier protein, sex hormone-binding globulin (SHBG), was significantly reduced. On the contrast, oxytocin is a non-lipid soluble hormone that plays a role in milk production in females. Consider if and how Oxytocin would be affected if its associated carrier protein, Neurophysin I, was significantly reduced. Hint: Consider the solubility, transport, and binding of each hormone
Endocrine System Signaling
The endocrine system uses 4 different types of hormone signaling to transmit information throughout the body. Autocrine signaling occurs when a cell secretes hormone molecules that bind to its own receptors. This sounds strange – why would a cell release a molecule that would bind to its own receptor? Releasing hormones that react on their own receptors allows cells to know the amount of hormone that they are releasing onto their environment. Paracrine signaling is when hormone molecules are secreted into the extracellular space and interact with nearby cells’ receptors. These hormones are secreted in small concentrations and are dependent on concentration gradients and diffusion for movement across the extracellular space.[3] Endocrine signaling is the most commonly known form of hormone signalling. Secreted hormones diffuse into either lymph vessels or the bloodstream and can reach distant tissues in the body.[4] This type of signaling often evokes more gradual and prolonged effects.[5] Lastly, neurocrine signaling occurs when nerves secrete chemical neurotransmitters into the extracellular space that can then diffuse into the bloodstream and interact with different tissues in the body. So like endocrine signaling, the chemicals travel down the bloodstream, but they are released from neurons as opposed to secretory cells. If you’re still having difficulty visualizing the different forms of communication, the diagram below visually summarizes the different types of hormone signaling within the endocrine system. Keep in mind that neurocrine signaling discussed in class is not displayed in this diagram.
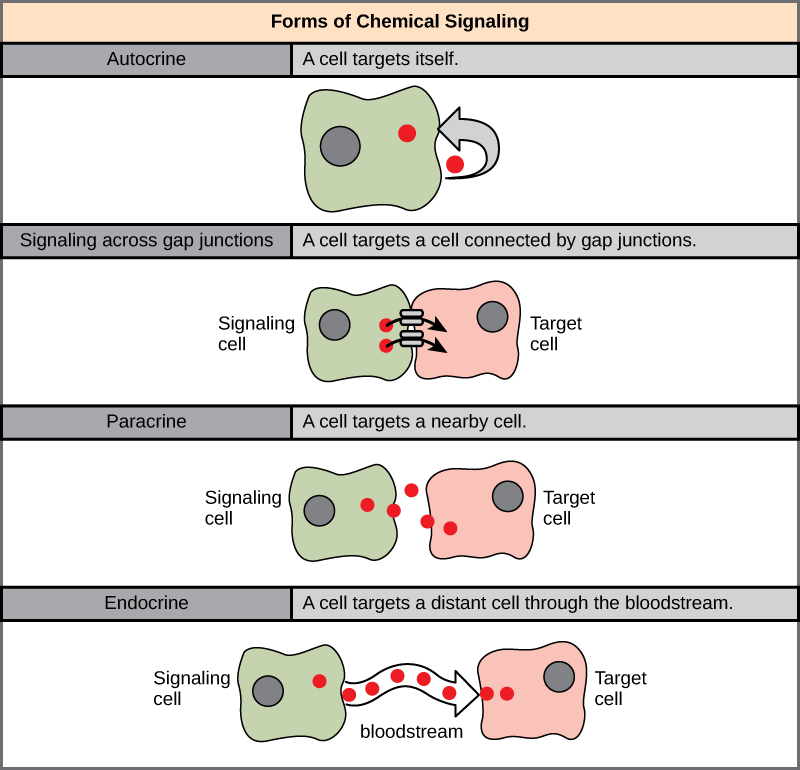
Test Your Knowledge
Real Life Scenario:
Let’s see how our knowledge of communication systems can be applied to the following scenario. Elevated levels of Carbon Dioxide or H+ in the blood will lead to an decrease in blood pH. Being a regulated variable, the body will act with immediate temporary compensations to raise the blood pH level. These short-term compensations will buy time until a proper correction can be made at the level of the kidneys which can take seconds to hours. In what circumstances would the body be using the nervous or endocrine system to compensate for disruptions in the environment? Hint: Consider the timing of communication responses.
Classes and Characteristics of Hormones
Classes of Hormones
The endocrine system uses a diverse means of communication within the body by using a combination of different classes of hormones to target specific receptors that elicit the appropriate response. Hormones are categorized into 3 main classes based on their structural and functional properties. The 3 main classes of hormones are peptides, amines and steroids.
Peptides – NLS Only
Peptide hormones are all NLS, meaning that they bind to their respective receptors only on cells’ plasma membranes. Peptide hormones, being NLS, can be stored in secretory vesicles of cells. This kind of storage saves secretory cells the hassle of synthesizing new hormones and allows them to be released “on-demand” for rapid action. Examples of peptide hormones include oxytocin, growth hormone, and follicle-stimulating hormone. Below is a visual representation of these hormones to help you differentiate peptide hormones from the other classes of hormones.
Amines (Metabolites of Amino Acids) – Both NLS and LS
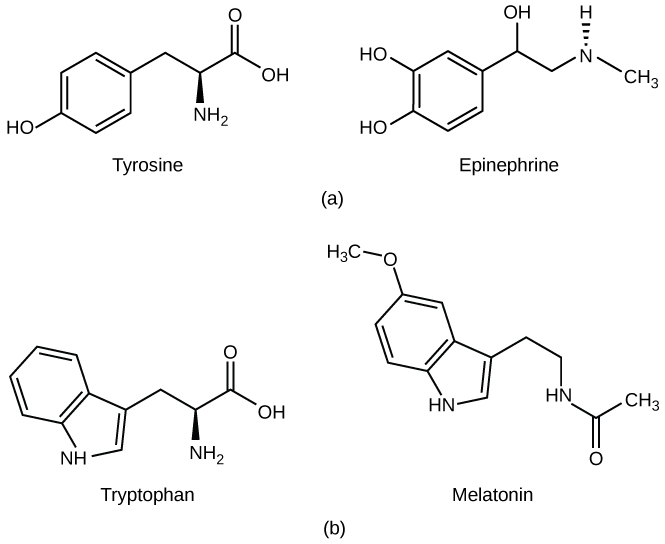
Amine hormones are derived from amino acids and can be either NLS or LS; however, they tend to be mostly NLS.[6]. NLS amine hormones bind to their respective receptors only on cells’ plasma membrane and can be stored in vesicles to be released “on-demand” for quicker response times. Meanwhile, LS amine hormones bind to their respective receptors either in the cytosol or in the nucleus. By interacting with receptors within the cell, this allows LS amine hormones to bypass signaling cascades to induce a more efficient cellular and genetic response. LS hormones cannot be stored within vesicles since they would be able to easily cross the vesicle membrane and constantly diffuse into the cytoplasm. However, very small concentrations of LS hormones can be stored bound to their carrier proteins; otherwise, they must be newly synthesized when needed. Examples of amine hormones include epinephrine, a NLS amine hormone, and melatonin, a LS amine hormone. Below is a visual representation of these hormones beside their respective amino acids of origin.
Steroids (Metabolites of Cholesterol) – LS
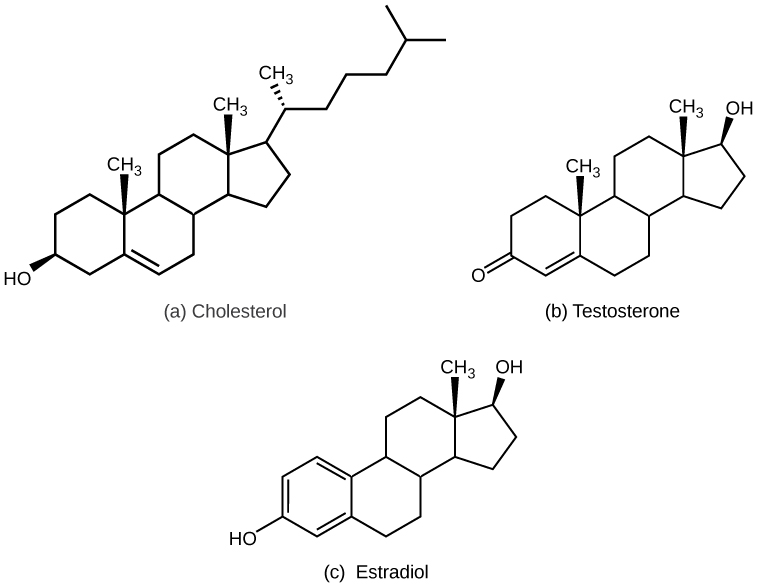
Steroid hormones are derived from cholesterol and are, almost essentially, all LS. This means steroid hormones bind to their respective receptors either in the cytosol or in the nucleus. Similar to LS amine hormones, steroid hormones cannot be stored within vesicles. As a result, steroid hormones must be either stored bound to their carrier protein or synthesized and secreted when needed. Examples of steroid hormones include testosterone and estradiol. Below is a visual representation of these hormones beside cholesterol from which they are derived. Take note of how structurally similar these hormones are to cholesterol.
Tips From Past Students
These visual examples of hormones are here just to help you understand the structural differences between the classes of hormones and why they would interact differently with their corresponding receptors – you are not expected to memorize these structures and you will never be tested on them in the HK*2810 course.
To help solidify our understanding of the classes of hormones, watch the following video that summarizes many of the key points regarding hormone-receptor actions. Keep in mind amine hormones are referred to in this video as amino acid derivatives. Additionally, although not discussed in this video, amine hormones can act through both nonsteroidal and steroidal hormone action since they can be NLS or LS.
“Mechanisms of Hormone Action” by Academic Algonquin is All Rights Reserved.
Tips From Past Students
Take your time and make sure that you fully understand these concepts as they are foundational to learning in-depth mechanisms of select hormones discussed later in this course.
General Characteristics of Hormones
Although hormones can be classified into 3 main classes based on their structural and functional properties, some characteristics are still shared across all classes of hormones. These general characteristics are described below.
Half-lives in Blood
All hormones, whether they’re amine, peptides, or steroids, have half-lives. This means that once they are released into the bloodstream they start to be degraded at a rate specific to the type of hormone. Once the hormones have completed their function and are no longer required, they can be eliminated from the body by the liver and kidneys.
Carrier Proteins
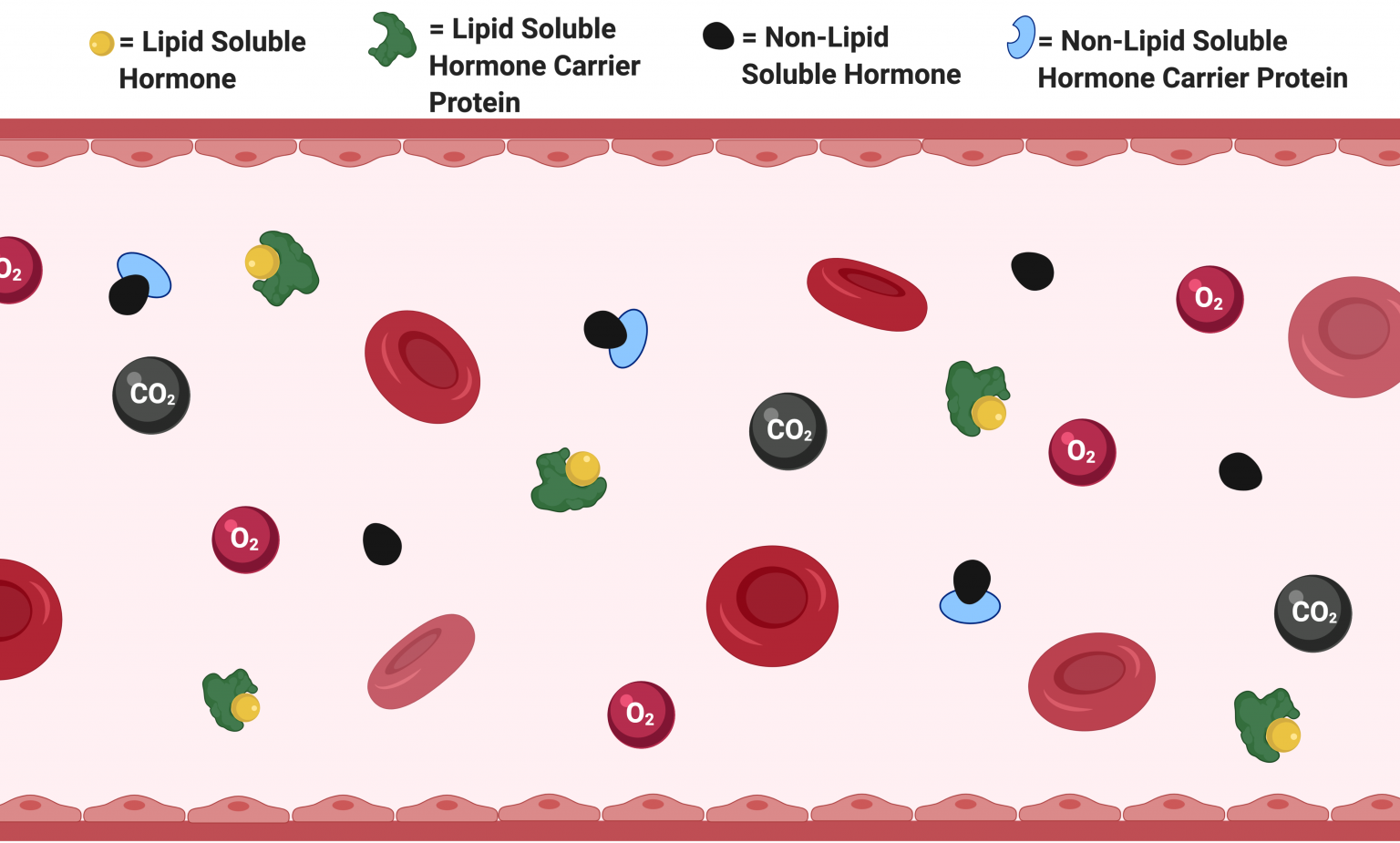
As mentioned previously, endocrine glands secrete hormones into the bloodstream where they dissolve into the plasma to be transported throughout the body to interact with their specific ligand receptors. Depending on the solubility of the hormone, it will travel through the bloodstream either free or bound to a carrier protein. Keep in mind that blood plasma consists mostly of water. As a result, LS hormone’s with their hydrophobic nature must be bound to a carrier protein to make them soluble enough for transport in the aqueous environment of the bloodstream. Meanwhile, NLS hormones with their hydrophilic nature exist in the bloodstream in an equilibrium between their free and carrier-bound state. However, as long as a hormone is bound to its carrier, it cannot interact with receptors. Therefore, in order to bind to target receptors, hormones have to be released from their carrier proteins.
In addition to facilitating transport, carrier proteins also act to provide a small reservoir of storage for hormones. A storage reservoir allows hormones to be released “on-demand” for rapid action rather than waiting for the hormone to be synthesized before taking an effect. In doing so, a storage reservoir acts to minimize fluctuations in hormone concentrations. Lastly, carrier proteins also function to extend hormones’ half-lives. Recall that hormones must be unbound from their carrier proteins before they can interact with their corresponding target receptors. Until unbound, a carrier-bound hormone will technically remain undetected in the body. As a result, by binding itself to a carrier protein, a hormone can extend its half-life in the body from minutes to days. Consequently, this makes hormone turnover much more efficient, as they remain stored and unused within the body for a prolonged period of time.
Receptors in Target Tissues
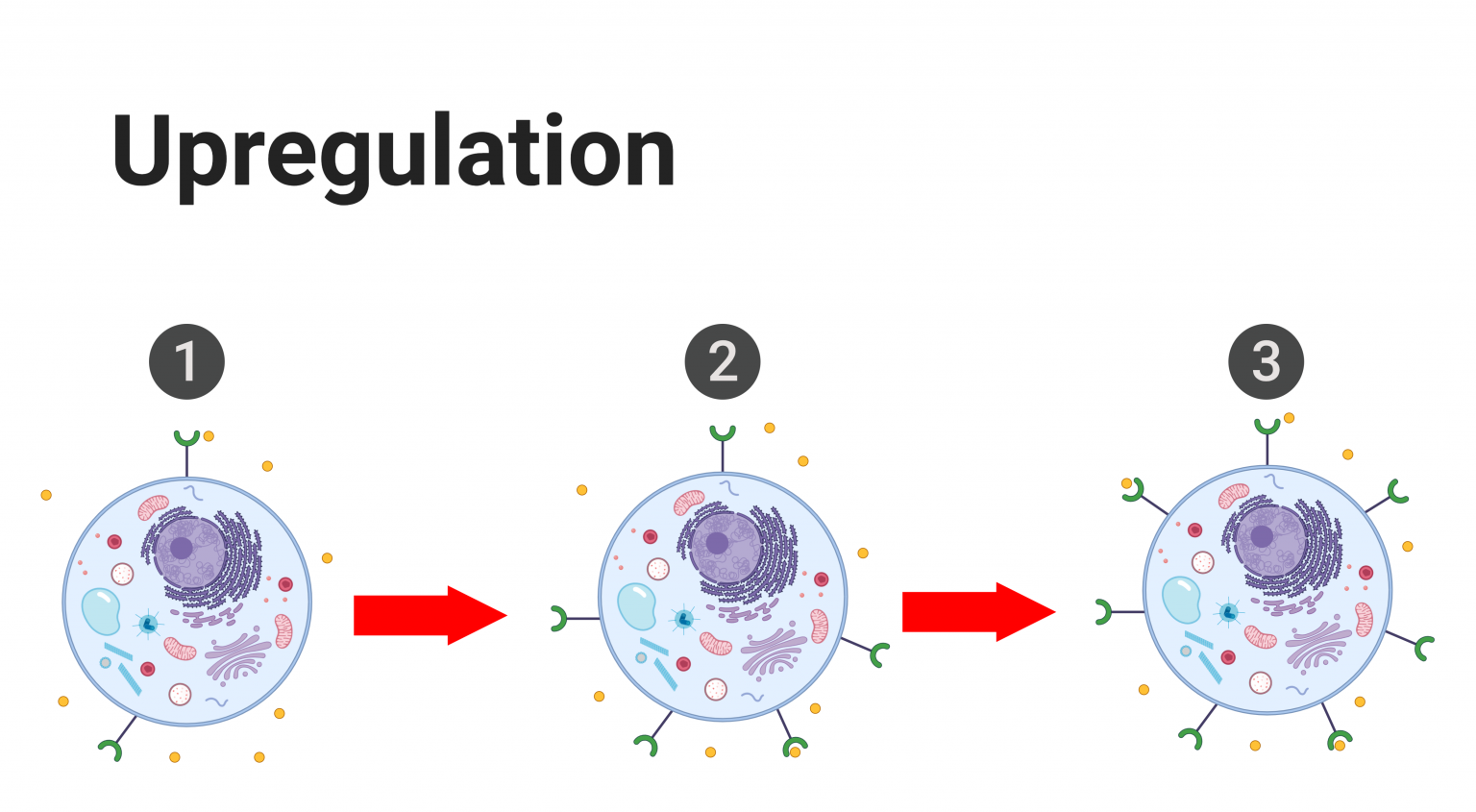
In order for a hormone to act on the body, two things need to happen: the hormone must be freed from its carrier protein and it must bind to a specific ligand-receptor on the target cell. Consequently, hormonal activity can be affected and regulated by altering the presence of hormone receptors on the target tissues. If a hormone’s circulating levels are low for a prolonged period of time (hours to days), cells can increase the number of target receptors to boost the body’s response to that respective hormone. This process is known as upregulation, and it increases the body’s sensitivity to the hormone since the hormone can now interact with more target receptors. On the other hand, if the levels of a given hormone are abnormally heightened, cells will decrease the number of target receptors to minimize the body’s response to that particular hormone when it is no longer needed. This process is known as downregulation, and it decreases the body’s sensitivity to the hormone.[7] Additionally, receptors, being proteins, are susceptible to damage and degradation. As a result, receptors are subject to constant turnover to meet the needs of the respective cell and the body. Therefore, the number of receptors on a plasma membrane or within a cell is never constant but always prone to fluctuations. Below is a visual representation of upregulation and downregulation. Keep in mind that upregulation and downregulation alter the concentration of hormone receptors on target cells, not hormones.
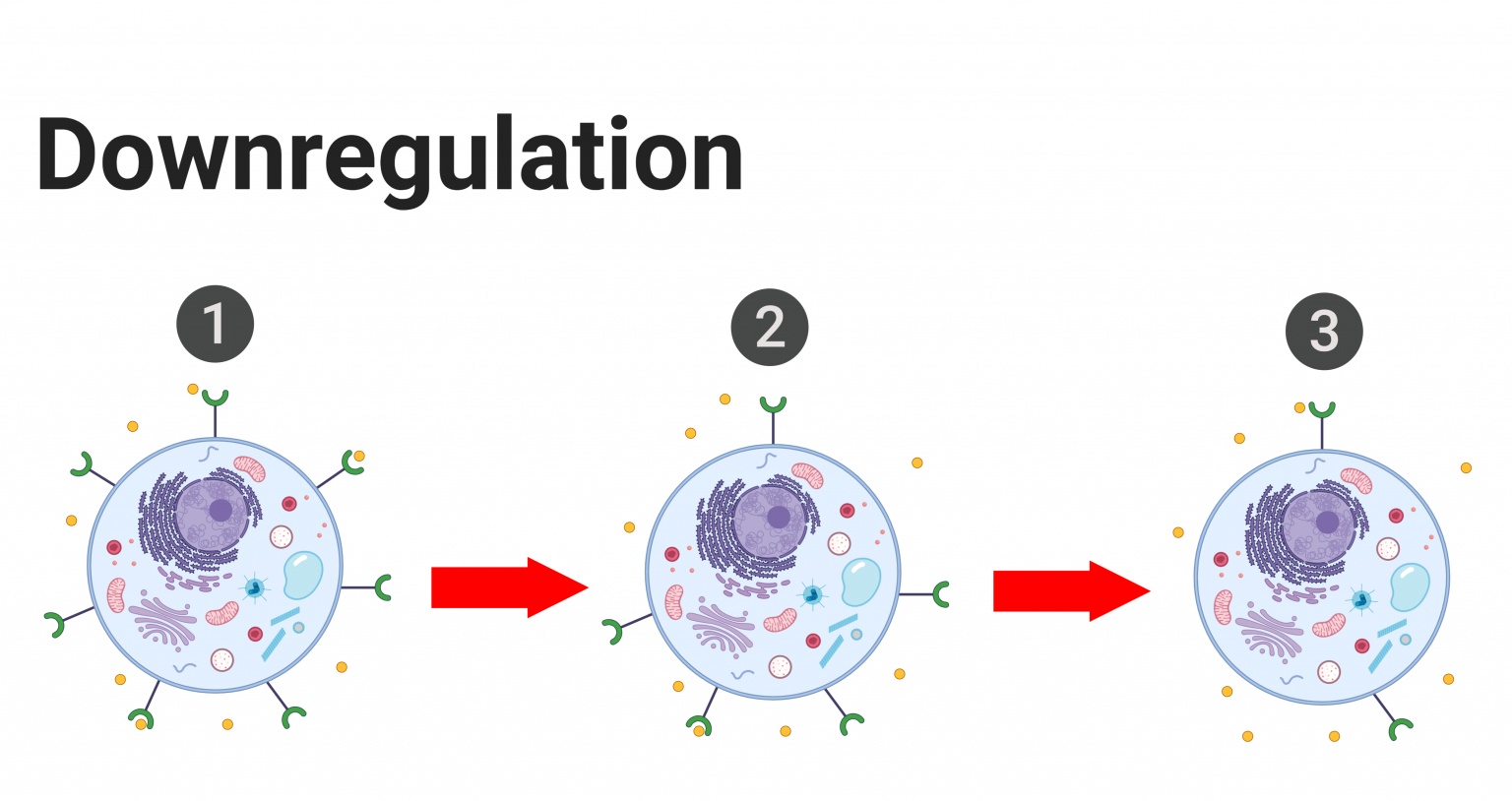
Tips From Past Students
Upregulation and downregulation only affect the body’s sensitivity to a given hormone by changing the concentration of target receptors. Upregulation and downregulation do not affect the concentration of hormones.
Permissive
Some hormones require the presence of another hormone in order to have its full effect or to function at all. An example discussed in class is the permissive relationship between the thyroid hormone and epinephrine. By itself, epinephrine has a weak effect on the release of free fatty acids from adipocytes. However, when secreted in the presence of thyroid hormone, epinephrine’s effect on adipocytes dramatically increases and it induces a massive release of free fatty acids from adipose tissue. This is different from additive signaling since thyroid hormone by itself does not affect the release of fat from adipocytes. Instead, the thyroid hormone functions by upregulating epinephrine receptors on the plasma membrane of target cells, and thus, increasing the body’s sensitivity and response to epinephrine.
Test Your Knowledge
Thinking Beyond:
Epinephrine is a hormone that can act at the level of arterioles to constrict vessels, therefore decreasing blood flow through the vessels. Ideally, epinephrine should not act on vessels near the heart to allow venous return. What is one structural feature that might prevent these vessels from constricting when epinephrine is released? Hint: Think about how hormones communicate with target cells.
Control of Hormone Secretion
Now that we have a foundational understanding of hormone signaling, classes, and general characteristics let’s explore how hormone secretion is controlled. Control of hormone secretion is essential to maintain homeostasis and coordinate bodily functions. Even a minor disturbance in hormonal control can lead to an imbalance which can have serious consequences on the body. There are three main forms of hormone secretion regulation: feedback loops, cyclic secretion, and neuronal regulation.
Feedback Loops
Out of the three mechanisms, feedback loops are the most common method of control. Feedback loops are tremendously important for the regulation of numerous physiological processes in the human body. Feedback loops can either be negative, which inhibits the overall release of the target hormone(s), or positive, which promotes the overall secretion of the target hormone(s).
Negative Feedback Loops
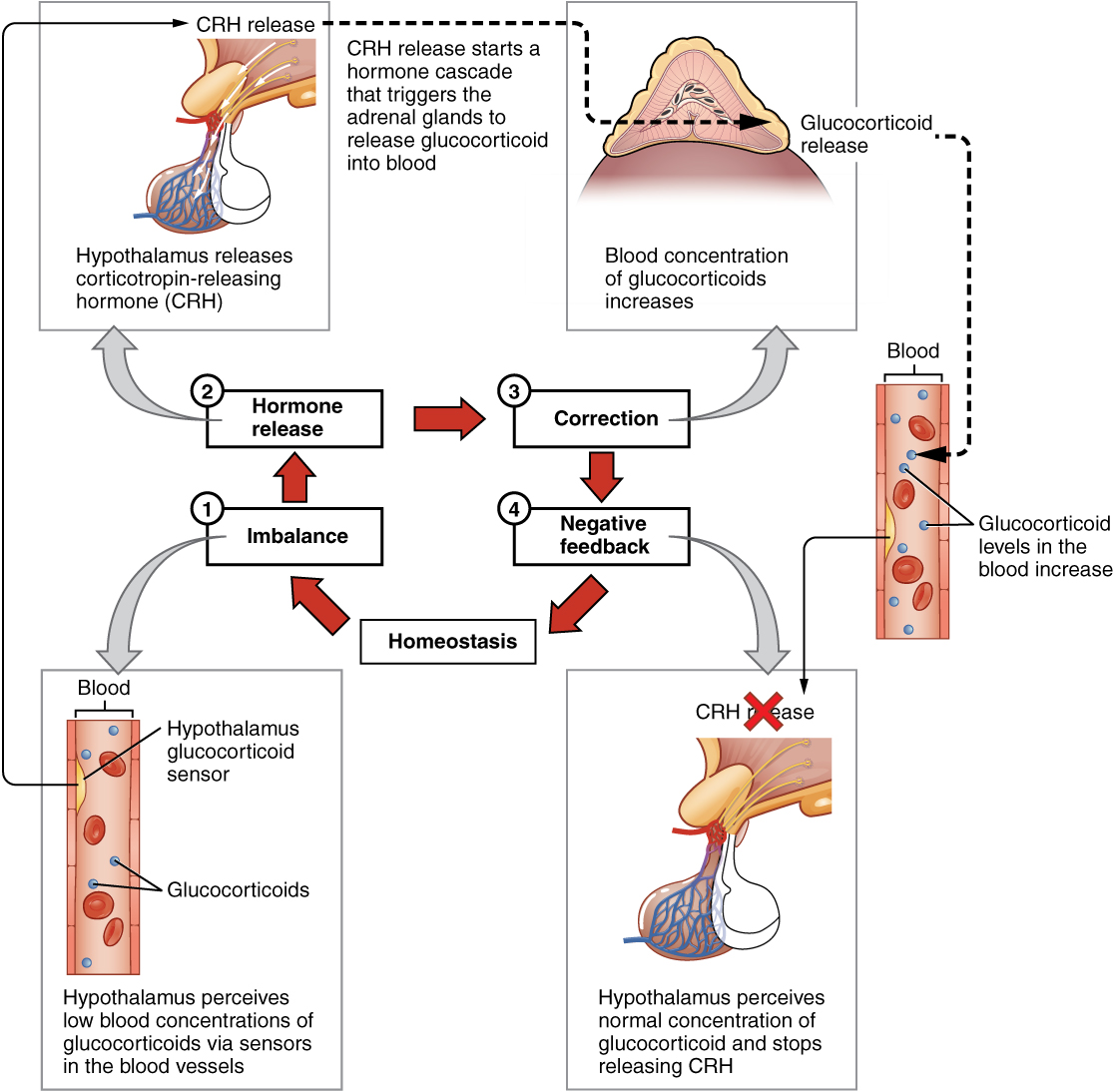
As discussed in class, negative feedback loops are used to regulate the homeostatic concentrations of hormones and regulated variables in order to maintain their appropriate ranges within the blood. In regards to hormone homeostasis, these loops respond to significant increases in hormone concentrations, and function by inhibiting different steps involved in the release of that corresponding hormone. Therefore, this type of loop regulation allows hormones to be tightly controlled within narrow ranges in the body. Below is an example of how the body regulates glucocorticoid blood concentrations. Glucocorticoids are a class of steroid hormone synthesized by the adrenal cortex most notably for its anti-inflammatory effects. Although glucocorticoid regulation is not covered or tested in this course, it makes use of regulation via a simple negative feedback loop. Subsequently, use the diagram below as a visual aid to help you understand the general concepts behind negative feedback loops.
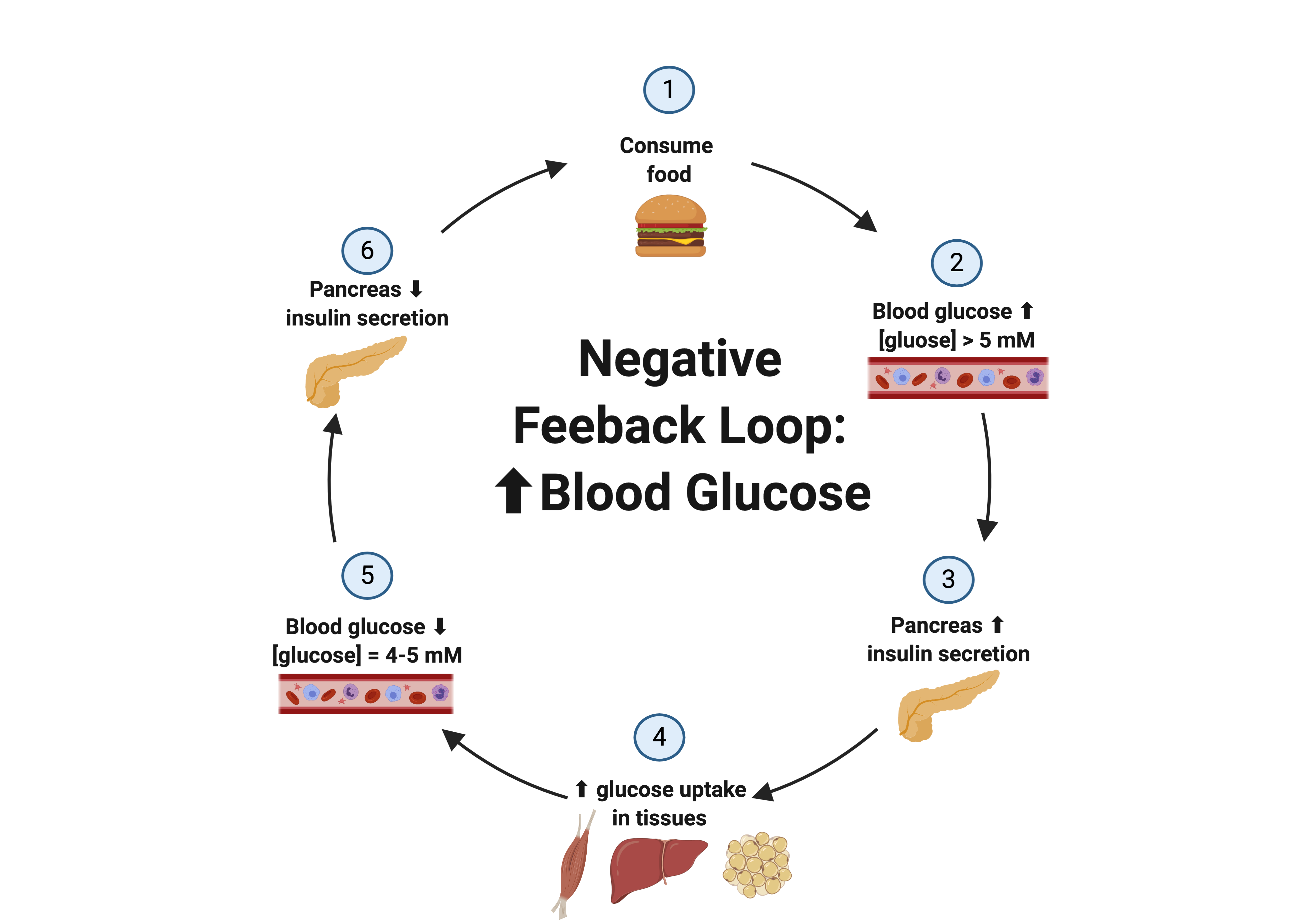
Negative feedback loops can also be used to manage regulated variables through changes in hormone concentrations. Take glucose regulation for example, as it will be discussed later in this chapter. Blood glucose levels are controlled within a tight range by using insulin and glucagon, two peptide hormones. If blood glucose levels are elevated, the body responds by secreting insulin from the pancreas. Insulin stimulates the uptake of glucose into tissues with insulin receptors, thus causing a decrease in blood glucose. Once blood glucose levels return to a normal range, insulin secretion is inhibited via a negative feedback loop. Below is a visual representation of the negative feedback loop in response to an increase in blood glucose levels.
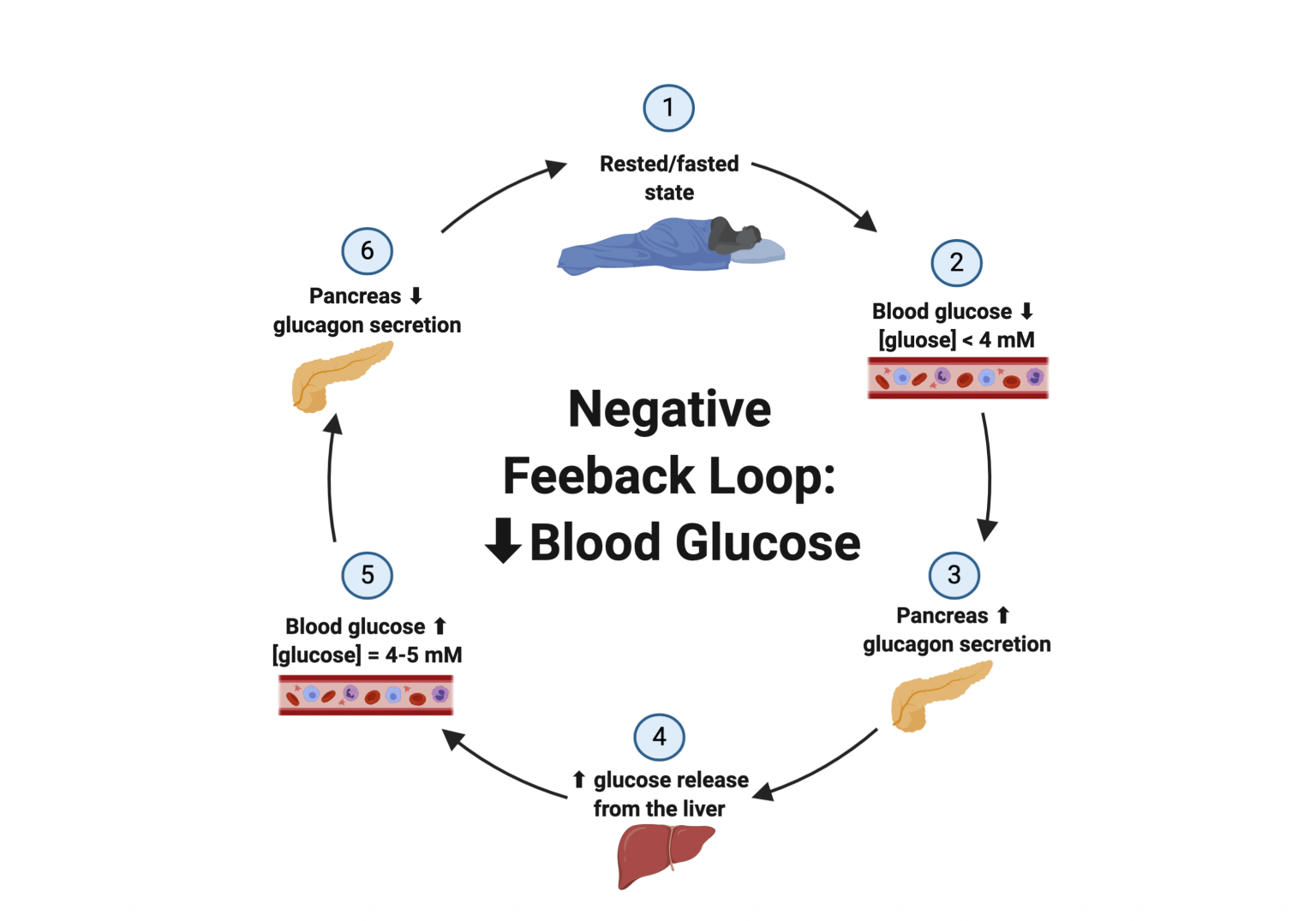
In the opposite sense, if blood glucose levels are reduced, the body responds by secreting glucagon from the pancreas. Glucagon stimulates the secretion of glucose from the liver, thus causing an increase in blood glucose. Once blood glucose levels return to a normal range, glucagon section is inhibited via a negative feedback loop. In this case, even though glucagon caused an increase in blood glucose levels, it is still considered a negative feedback loop since the increase in blood glucose signaled the inhibition of glucagon secretion. Below is a visual representation of the negative feedback loop in response to a decrease in blood glucose levels.
Negative feedback can be thought of as regulating “minor” everyday shifts to homeostases, such as changes in blood glucose and body temperature. Meanwhile, positive feedback is used for more “serious” situations that require amplification of our body’s response. Additionally, instead of bringing a variable back to its physiological set point, a positive feedback loop will further increase its deviation from homeostasis. It is also inportant to be aware that the examples of negative feedback loops provided above are very simple in the sense that hormone secretion is only inhibited at one step. In reality, negative feedback loops can be quite complex and can make use of various hormones and involve inhibition at multiple steps within a pathway. These simplistic examples were provided to help you nail down the concepts of negative feedback since it is an essential foundation for this course and future physiology courses. For the moment, you are not required to know the in-depth mechanisms of the hormonal regulation of glucocorticoids or glucose; however, be sure to fully understand the general concepts behind negative feedback loops.
Positive Feedback Loops
While negative feedback loops inhibit the further secretion of a hormone, positive feedback loops promote the continual release of the hormone. In a positive feedback loop, the release of a certain hormone will trigger a pathway that will continue to encourage its release to increase the hormone’s concentration beyond the normal homeostatic range. This process will continue to increase the hormone levels and augment its effects until an external stimulus signals the body to stop. This amplification of hormone secretion is quite powerful and its use is mostly reserved for situations when our lives are “at-risk”. Although greatly beneficial and essential to our survival, if left unregulated, this loop could be problematic and cause adverse consequences.
A well-known example of a positive feedback loop is the release of oxytocin from the posterior pituitary gland during childbirth. During childbirth, the baby will stretch the wall of the cervix which will stimulate the release of oxytocin. Oxytocin acts to strengthen the contractions that push the baby out of the mother’s womb. As the baby’s delivery progresses, the cervix will continue to stretch, stimulating even more oxytocin release, and further intensify the contractions. Once the baby is fully delivered, the cervix will no longer be stretched which is the external stimulus to cease oxytocin release thus, ending the positive feedback loop. Below is a visual representation of the positive feedback loop of oxytocin release in response to childbirth.
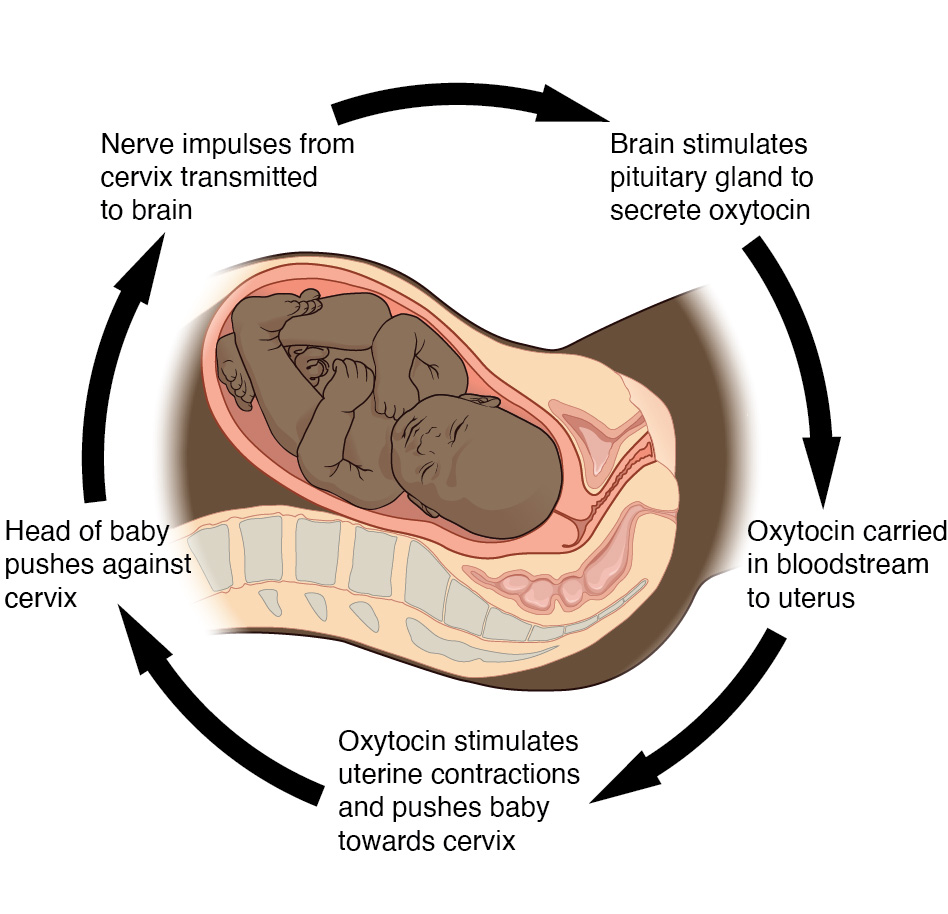
Tips From Past Students
For the moment, you are not required to know the in-depth mechanisms of the oxytocin release in response to childbirth; however, be sure to fully understand the general concepts behind negative and positive feedback loops.
To help you differentiate between positive and negative feedback loops, the video below explains both examples discussed in this chapter, glucose regulation, and childbirth. The video also illustrates additional feedback loops discussed briefly in class, such as body temperature regulation (a negative feedback loop) and blood clotting (a positive feedback loop). At this point in the course, do not worry about memorizing details of the different pathways described in this video, focus on the general concepts behind negative and positive feedback loops.
“Negative vs Positive Feedback” by Beverly Biology is All Rights Reserved.
Cyclic Secretion
Cyclic secretions are used when the body wants to produce and secrete a hormone at defined times and without requiring specific external stimuli. This defined time can be daily, monthly, seasonal, or even during specific developmental periods. This specific type of hormone regulation occurs in the suprachiasmatic nucleus of the hypothalamus, which controls our circadian clock. In order to regulate the production and secretion of hormones cyclically, the suprachiasmatic nucleus uses information from different sources, such as the light-dark cycle and the body’s sleep-wake cycle. This integration of information allows the hypothalamus to predict human behavior and coordinate appropriate rhythms for endocrine, metabolic, and behavioral activity. An example of this level of regulation is the daily secretion of growth hormone (GH) and adrenocorticotropic hormone (ACTH). These two hormones are released at specific periods during our sleep cycle and their scheduled release is important to obtain a good night’s rest. The graph below is to help you understand the importance of cyclic secretion and appreciate how it allows for the production and release of certain hormones at specific times. You are not expected to know any values related to these two hormones but rather appreciate the general concepts behind cyclic secretion.
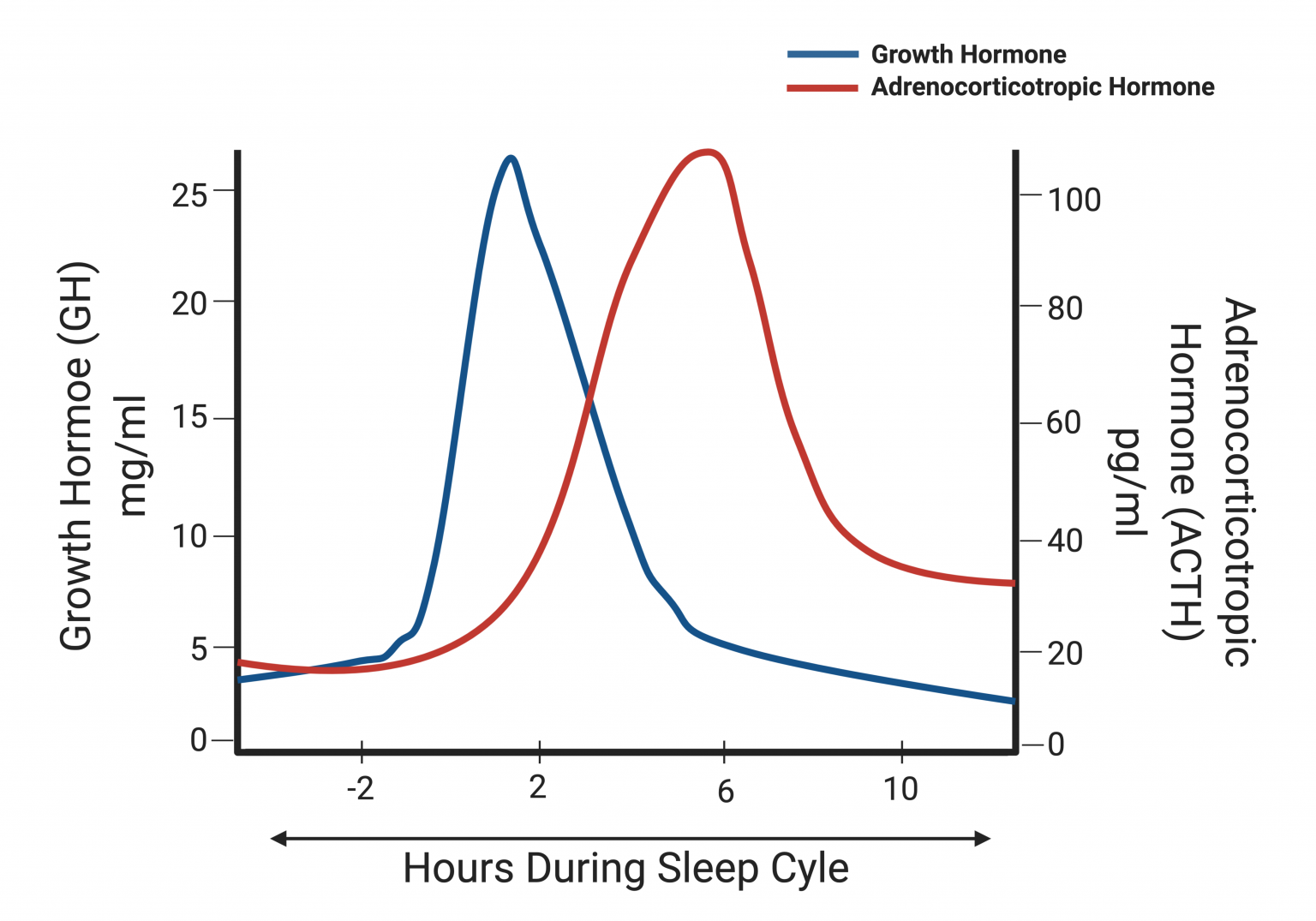
Test Your Knowledge
Real-Life Scenario:
Melatonin is a hormone that is secreted in the body to aid in sleep. What is keeping this hormone from being released in the middle of the day, and being released when you need to sleep? Describe this process? Hint: Consider the type of hormonal control that occurs at regular intervals.
Examples of other hormones regulated in a cyclic manner include female sex hormones which will be discussed in more depth later in this chapter. Female sex hormones such as luteinizing hormone (LH) and follicle-stimulating hormone (FSH), are secreted more gradually over a period of days as opposed to hours. Below is a diagram displaying the cyclic secretion of four sex hormones during a woman’s menstrual cycle. At this point in the course, do not worry about memorizing specific details regarding the different hormone levels throughout the menstrual cycle, rather focus on the general concepts behind cyclic secretion.
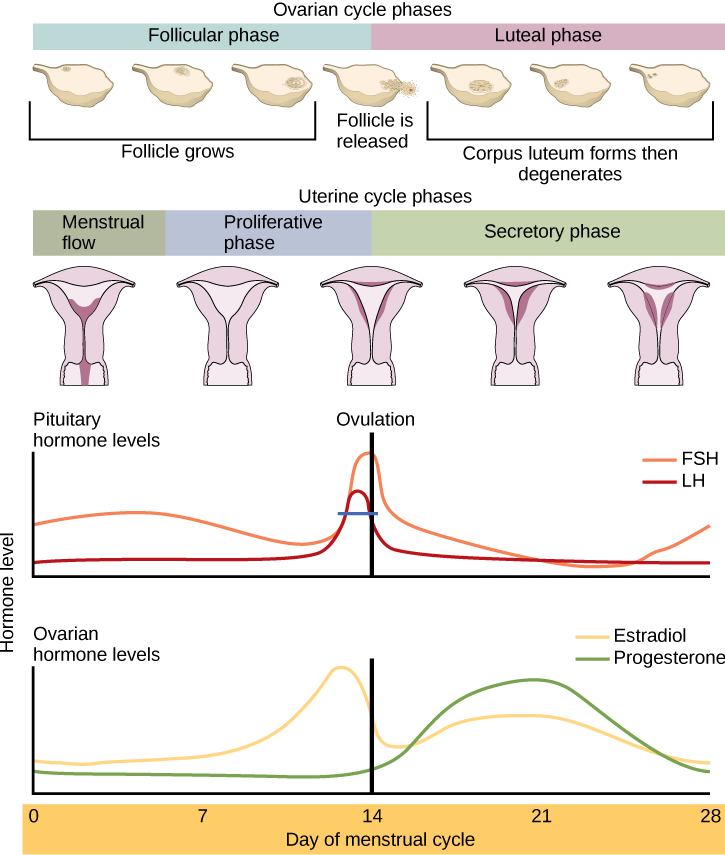
Tips From Past Students
These diagrams are here just to help you understand how vast the periods of cyclic secretion can be. Some hormones’ levels rise and fall each day, while other hormones are regulated at a monthly time period. Other hormones even experience cyclic regulation over a period of years, such as growth hormones and sex hormones during puberty.
Neuronal
The central nervous system can also regulate hormone secretion. This type of regulation will not be discussed in depth in this chapter and can be further explored in the Hypothalamus and Pituitary subchapter.
Key Takeaways
- Compare and contrast between the two communication systems, nervous and endocrine.
- How lipid-soluble and non-lipid soluble hormones function within the body.
- How carrier proteins help hormones interact with their target receptors to elicit a response.
- Why and how target receptors are upregulated and downregulated.
- Understand the general concepts behind the types of control over hormone secretion.
Subchapter Quiz
The questions below can be used to assess your knowledge within this chapter. There are five multiple-choice questions that you should attempt without referring to your notes. The questions will provide you with responses to your answers to guide your studying but should not be used as your only resource.
Media Attributions
- Binding of Water Soluble Hormones © OpenStax is licensed under a CC BY (Attribution) license
- Binding of Lipid-Soluble Hormones © OpenStax is licensed under a CC BY (Attribution) license
- Forms of Chemical Signaling © OpenStax is licensed under a CC BY (Attribution) license
- Examples of Peptide Hormones © OpenStax is licensed under a CC BY (Attribution) license
- Examples of Amine Hormones © OpenStax is licensed under a CC BY (Attribution) license
- Examples of Steroid Hormones © OpenStax is licensed under a CC BY (Attribution) license
- Lipid-soluble and non-lipid soluble hormones travelling in the bloodstream © Daniel Barros is licensed under a All Rights Reserved license
- Upregulation © Daniel Barros is licensed under a All Rights Reserved license
- Downregulation © Daniel Barros is licensed under a All Rights Reserved license
- Glucocorticoid Regulation © OpenStax is licensed under a CC BY (Attribution) license
- Negative Feedback Loop: Increased Blood Glucose © Daniel Barros is licensed under a All Rights Reserved license
- Negative Feedback Loop: Decreased Blood Glucose © Daniel Barros is licensed under a All Rights Reserved license
- Positive Feedback: Oxytocin © OpenStax is licensed under a CC BY (Attribution) license
- Cyclic Secretion: Growth Hormone (GH) and Adrenocorticotropic Hormone (ACTH) © Daniel Barros is licensed under a All Rights Reserved license
- Cyclic Secretion: Ovarian Cycle © OpenStax is licensed under a CC BY (Attribution) license
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013). An Overview of the Endocrine System. Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013). An Overview of the Endocrine System. Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013).Signaling Molecules and Cellular Receptors.Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013). An Overview of the Endocrine System. Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013).Signaling Molecules and Cellular Receptors.Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013).Types of Hormones. Anatomy and Physiology. ↵
- Betts, G. J., Young, K. A., Wise, J. A., Johnson, W., Poe, B., Kruse, DH., Korol, O., Johnson, J. E., Womble, M., & DeSaix, P. (2013).Types of Hormones. Anatomy and Physiology. ↵
A state of relatively stable equilibrium in the body that is maintained by constant adjustment of biochemical and physiological pathways.
The network of nerve cells and fibers which transmit nerve impulses between parts of the body.
Chemical released at synapses by nerves in response to a stimulus.
System of glands that produce hormones to regulate and control bodily activities.
Signalling chemicals secreted by endocrine cells. These chemicals may be released onto the extracellular space or the bloodstream.
Cell signalling in which a cell secretes hormone molecules that binds to receptors on that same cell.
A form of cellular signalling in which cells that are near one another communicate through the release of chemical messengers, allowing cells to locally coordinate their activities.
Cell signalling in which a cell secretes hormones that diffuse into either lymph vessels or the bloodstream to reach distant cells in the body.
Cell signalling in which nerves secrete chemical neurotransmitters into the extracellular space that diffuse into the bloodstream and interact with different tissues in the body.
The time it takes for hormones to lose half of its activity by degradation or elimination.
A type of cell membrane protein involved in facilitated diffusion and active transport of substances across the membrane.
An increase in the production of a target receptor that causes an increase in the body's sensitivity to a particular hormone.
A decrease in the production of a target receptor that, in turn, causes a decrease in the body's sensitivity to a particular hormone.
A hormone that requires the presence of an additional hormone to have its full effect in the body; the additional hormone by itself will have no effect.
The presence of a hormone will further increase the effect of an already acting hormone by producing an effect of similar nature; both of these hormones can function on their own.
Occurs when some function of the output of a system, process, or mechanism, is fed back in a manner that tends to reduce the output.
A variable that is being sensed and maintained by a homeostatic system.
Occurs when some function of the output of a system, process, or mechanism is fed back in a manner that tends to increase the output.
Small portion of the brain that regulates metabolic processes, serving as a link between endocrine and nervous systems via the pituitary gland.
