
Eleutherococcus sessiliflorus Inhibits Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-Induced Osteoclast Differentiation and Prevents Ovariectomy (OVX)-Induced Bone Loss


Abstract
:1. Introduction
2. Results
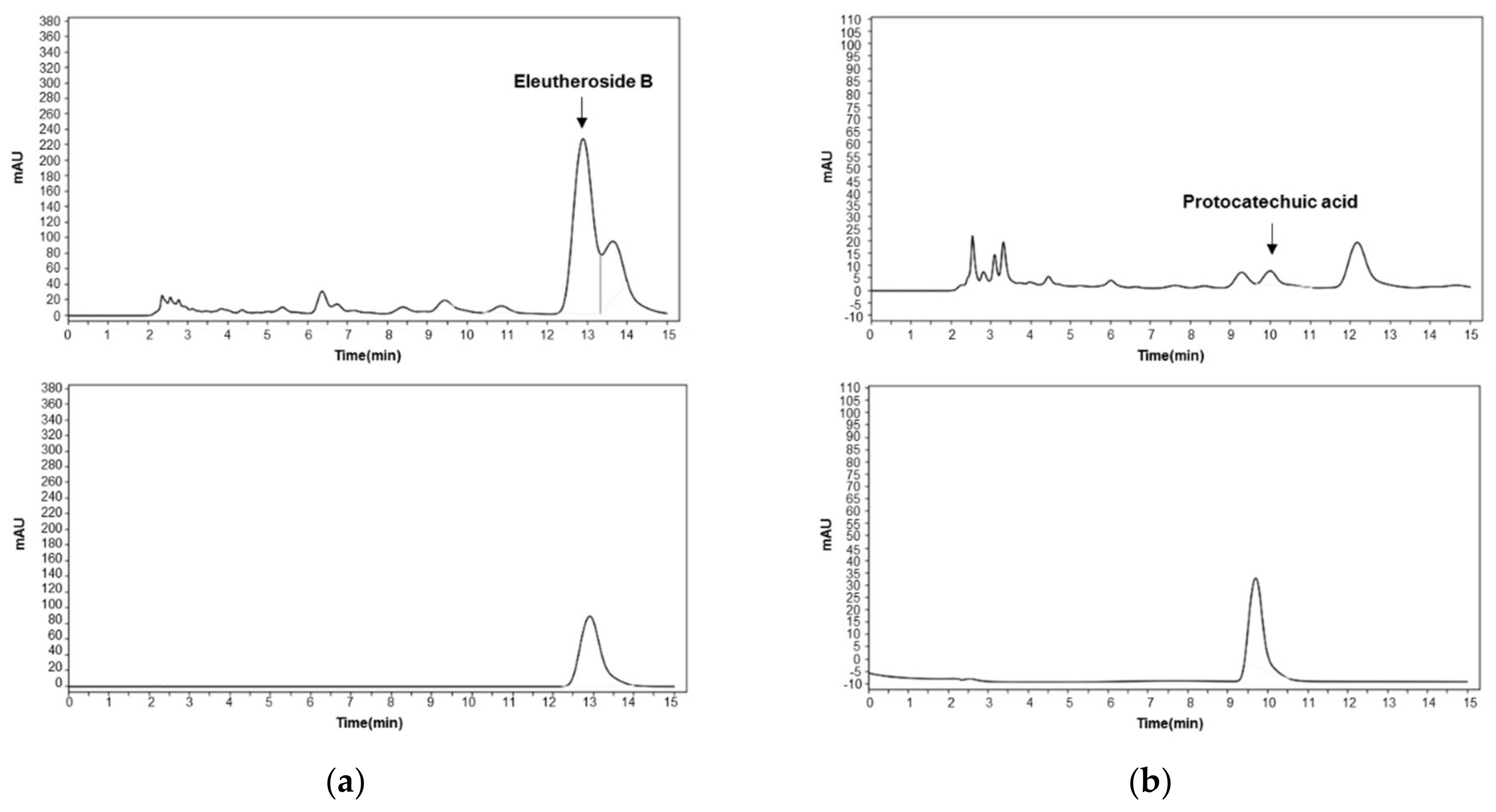
2.1. High-Performance Liquid Chromatography (HPLC) Analysis of ES Water Extracts
2.2. ES Inhibits RANKL-Induced Osteoclastogenesis
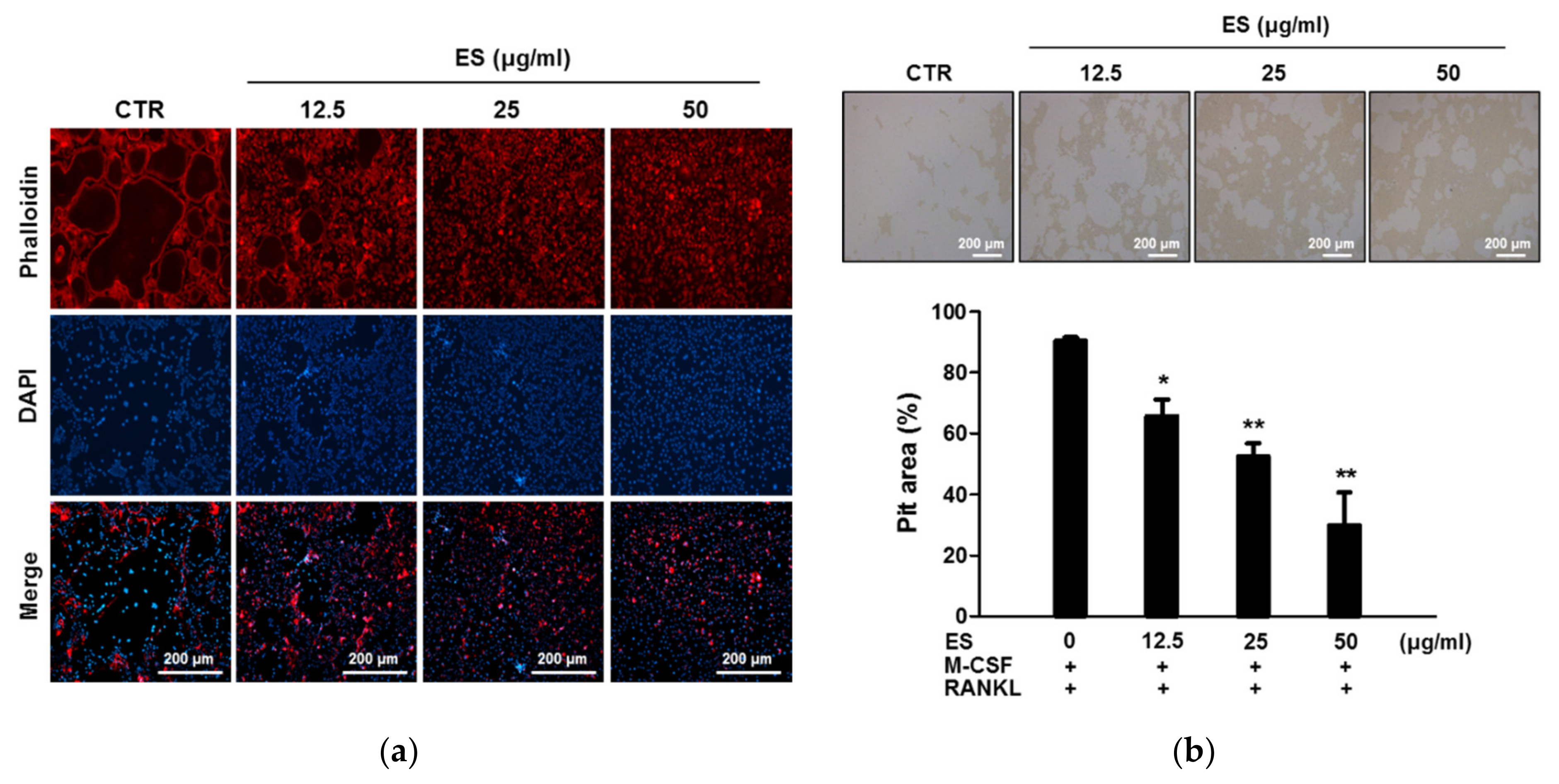
2.3. ES Inhibits Actin Belts Formation and Bone Resorption in Osteoclasts
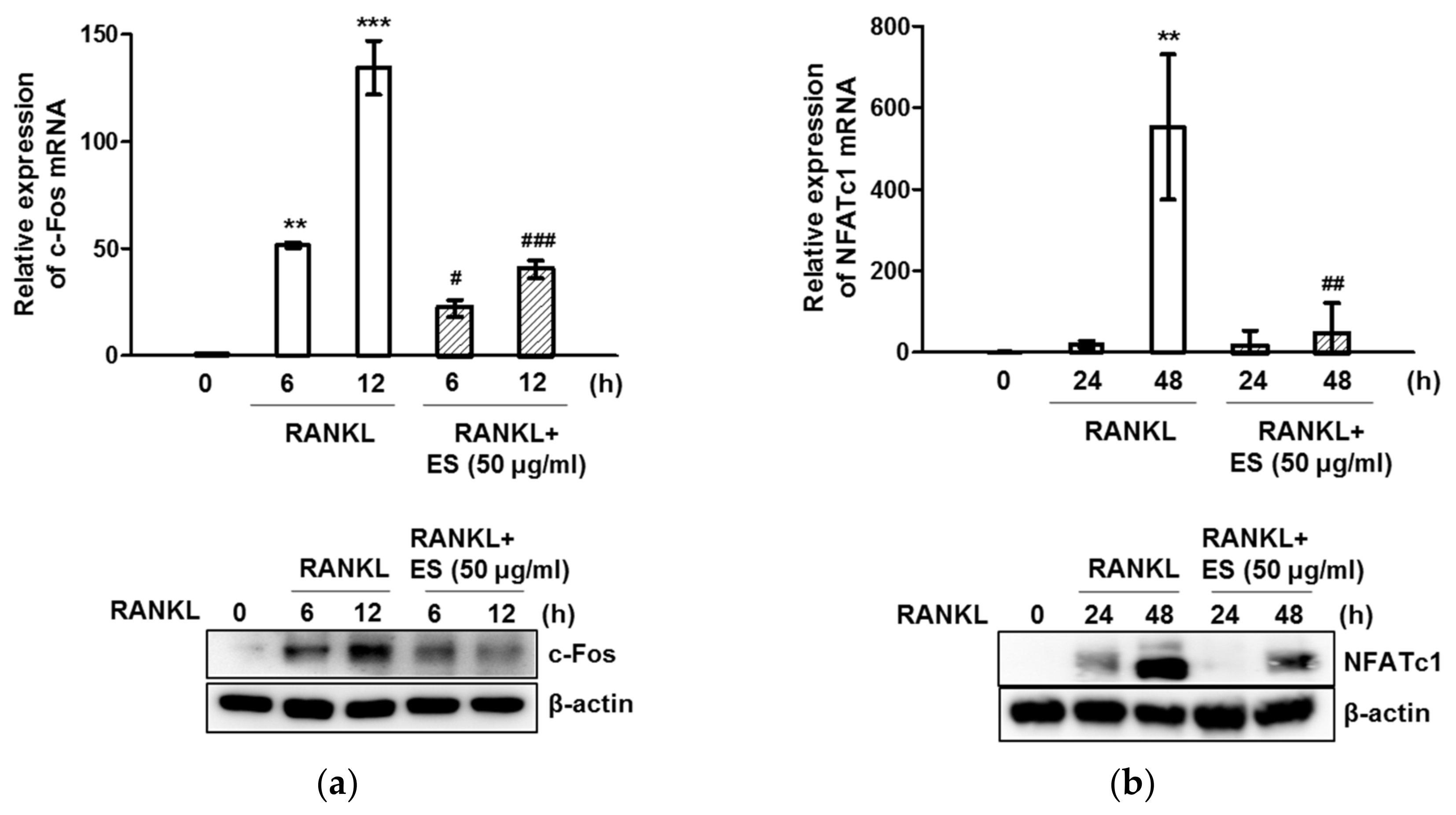
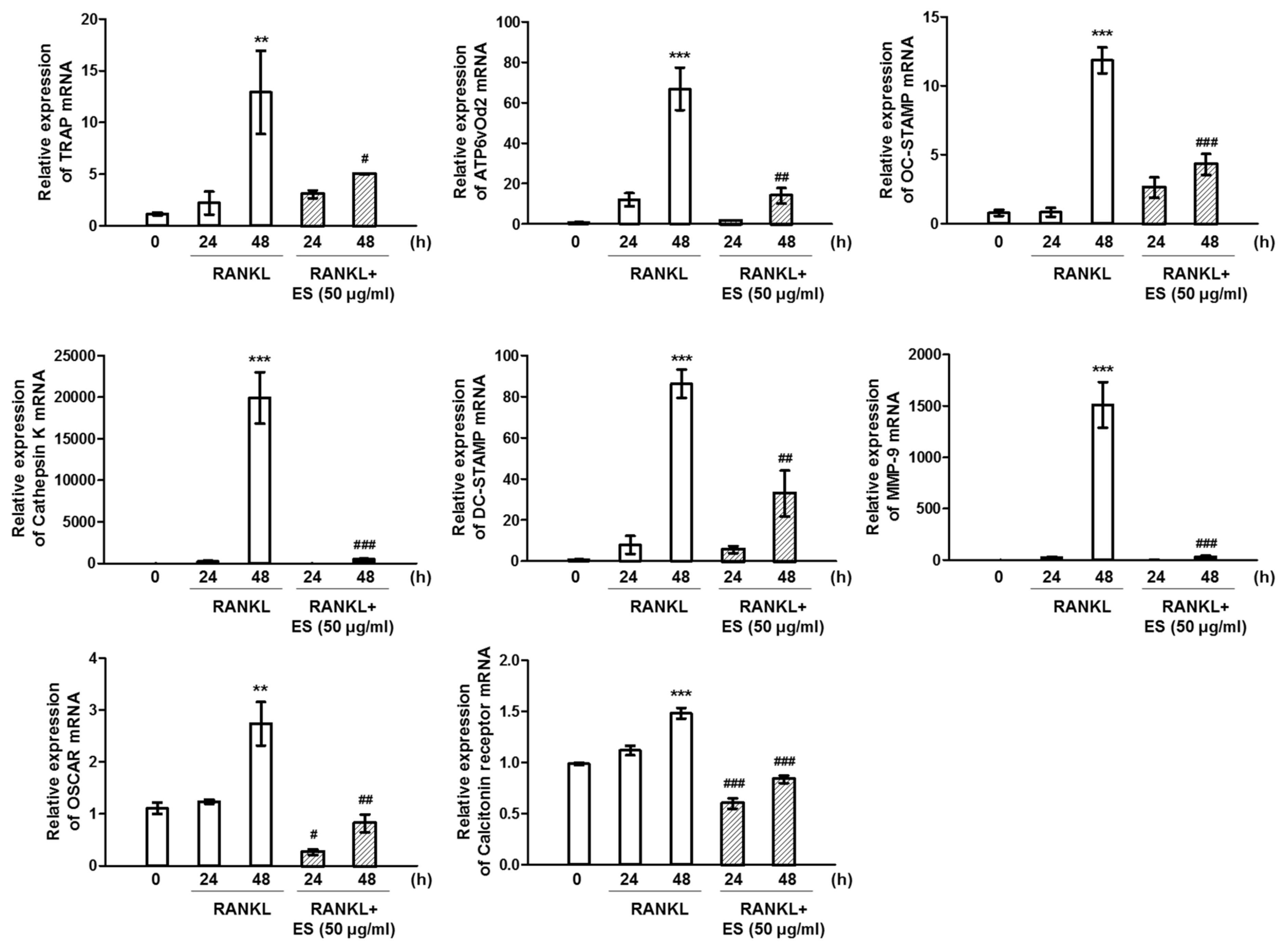
2.4. ES inhibits RANKL-Induced c-Fos and NFATc1 and Osteoclast-Specific Gene Expression
2.5. ES Inhibits Osteoclastogenesis via MAPK and NF-κB Signaling
2.6. ES Attenuates OVX-Mediated Bone Loss
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of ES
4.3. HPLC Analysis
4.4. Isolation of BMMs
4.5. Osteoclast Differentiation and TRAP Assay
4.6. F-actin Belts Staining
4.7. Cell Viability
4.8. Bone Resorption Assay
4.9. Western Blot Analysis
4.10. Real-Time RT-PCR Analysis
4.11. Animal Bone Loss Model
4.12. Micro-CT and Histological Analysis
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Borgström, F.; Karlsson, L.; Ortsäter, G.; Norton, N.; Halbout, P.; Cooper, C.; Lorentzon, M.; McCloskey, E.V.; Harvey, N.C.; Javaid, M.K.; et al. Fragility fractures in Europe: Burden, management and opportunities. Arch. Osteoporos. 2020, 15, 59. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.M.; Russell, L.; Khan, S.N. Osteoporosis. Clin. Orthop. Relat. Res. 2000, 372, 139–150. [Google Scholar] [CrossRef]
- Chen, L.R.; Ko, N.Y.; Chen, K.H. Medical treatment for osteoporosis: From molecular to clinical opinions. Int. J. Mol. Sci. 2019, 20, 2213. [Google Scholar] [CrossRef] [Green Version]
- Russow, G.; Jahn, D.; Appelt, J.; Märdian, S.; Tsitsilonis, S.; Keller, J. Anabolic Therapies in Osteoporosis and Bone Regeneration. Int. J. Mol. Sci. 2018, 20, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, D.H.; Rekedal, L.; Cadarette, S.M. Osteoporosis treatments and adverse events. Curr. Opin. Rheumatol. 2009, 21, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Kang, K.S.; Chun, K.H.; Hwang, G.S. Cordyceps militaris mushroom and cordycepin inhibit RANKL-induced osteoclast differentiation. J. Med. Food 2015, 18, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.T.; Dang, N.H.; Kim, O.; Van Cuong, P.; Dat, N.T.; Hwangbo, C.; Van Minh, C.; Lee, J.H. Ethanol extract of Polyscias fruticosa leaves suppresses RANKL-mediated osteoclastogenesis in vitro and LPS-induced bone loss in vivo. Phytomed. Int. J. Phytother. Phytopharm. 2019, 59, 152908. [Google Scholar] [CrossRef] [PubMed]
- Rho, T.W.; Lee, S.Y.; Han, S.Y.; Kim, J.H.; Lee, K.H.; Kim, D.S.; Kwak, H.B.; Kim, Y.K. Glycyrrhizae radix inhibits osteoclast differentiation by inhibiting c-Fos-dependent NFATc1 expression. Am. J. Chin. Med. 2017, 45, 283–298. [Google Scholar] [CrossRef]
- Hwang, J.H.; Cha, P.H.; Han, G.; Bach, T.T.; Min, D.S.; Choi, K.Y. Euodia sutchuenensis Dode extract stimulates osteoblast differentiation via Wnt/β-catenin pathway activation. Exp. Mol. Med. 2015, 47, e152. [Google Scholar] [CrossRef] [Green Version]
- Tasadduq, R.; Gordon, J.; Al-Ghanim, K.A.; Lian, J.B.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Shakoori, A.R. Ethanol extract of cissus quadrangularis enhances osteoblast differentiation and mineralization of murine pre-osteoblastic MC3T3-E1 cells. J. Cell. Physiol. 2017, 232, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Bensky, D.; Gamble, A. Chinese Herbal Medicine Materia Medica, 3rd ed.; Eastland Press: Seattle, WA, USA, 2004; pp. 338–340. [Google Scholar]
- Shikov, A.N.; Narkevich, I.A.; Flisyuk, E.V.; Luzhanin, V.G.; Pozharitskaya, O.N. Medicinal plants from the 14(th) edition of the Russian Pharmacopoeia, recent updates. J. Ethnopharmacol. 2021, 268, 113685. [Google Scholar] [CrossRef]
- Korea Food and Drug Administration (KFDA). Korean Pharmacopoeia, 9th ed.; KFDA: Seoul, Korea, 2007; p. 76. [Google Scholar]
- China Food and Drug Administration (CFDA). People’s Republic of China Pharmacopoeia, 10th ed.; China Medical Science Press: Beijing, China, 2015; pp. 66–206. [Google Scholar]
- European Directorate for the quality of Medicines & Healthcare. European Pharmacopoeia, 9th ed.; European Pharmacopoeia: Strasbourg, France, 2017; pp. 1230, 1344–1346. [Google Scholar]
- United States Pharmacopeial Convention. United States Pharmacopeia Dietary Supplements Compendium; United States Pharmacopeial Convention: Rockville, MD, USA, 2015; p. 1085. [Google Scholar]
- Jung, S.K.; Lee, H.J.; Lee, K.W. Bioactivities and action mechanisms of Acanthopanax species. Food Sci. Biotechnol. 2012, 21, 1227–1233. [Google Scholar] [CrossRef]
- Song, Y.; Deng, Y.; Huang, D.; Wen, J.; Liu, Z.; Li, F. LC-MS/MS determination and pharmacokinetic study of four lignan components in rat plasma after oral administration of Acanthopanax sessiliflorus extract. J. Ethnopharmacol. 2012, 141, 957–963. [Google Scholar] [CrossRef]
- Seo, B.I.; Kwon, D.Y.; Choi, H.Y.; Lee, H.J.; Oh, M.S.; Bu, Y.M. Medicinal Herbology; Younglim-Sa: Seoul, Korea, 2015; pp. 382–383. [Google Scholar]
- Kim, M.J.; Wang, H.S.; Lee, M.W. Anti-inflammatory effects of fermented bark of Acanthopanax sessiliflorus and its isolated compounds on lipopolysaccharide-treated RAW 264.7 macrophage cells. Evid. Based Complementary Altern. Med. 2020, 2020, 6749425. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, J.; Liu, M.; Li, Y.; Pan, J.; Liu, H.; Wang, W.; Bai, D.; Xiang, L.; Xiao, G.G.; et al. Therapeutic effects of Cortex acanthopanacis Aqueous extract on bone metabolism of ovariectomized rats. Evid. Based Complementary Altern. Med. 2012, 2012, 492627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, B.E.; Li, Q.X.; Liang, W.J. Experimental study on anti-tumor effects of cortex Acanthopanacis senticosus in vivo and in vitro. Chin. J. Integr. Tradit. West. Med. 2004, 24, 55–58. [Google Scholar]
- Thamizhiniyan, V.; Young-Woong, C.; Young-Kyoon, K. The cytotoxic nature of Acanthopanax sessiliflorus stem bark extracts in human breast cancer cells. Saudi J. Biol. Sci. 2015, 22, 752–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.K.; Kim, C.K.; Gong, S.K.; Yu, A.R.; Lee, M.Y.; Park, S.K. Acanthopanax sessiliflorus stem confers increased resistance to environmental stresses and lifespan extension in Caenorhabditis elegans. Nutr. Res. Pract. 2014, 8, 526–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bari, A.A.; Al Mamun, A. Current advances in regulation of bone homeostasis. FASEB Bioadv. 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Feng, X.; Teitelbaum, S.L. Osteoclasts: New insights. Bone Res. 2013, 1, 11–26. [Google Scholar]
- Wiktor-Jedrzejczak, W.; Bartocci, A.; Ferrante, A.W., Jr.; Ahmed-Ansari, A.; Sell, K.W.; Pollard, J.W.; Stanley, E.R. Total absence of colony-stimulating factor 1 in the macrophage-deficient osteopetrotic (op/op) mouse. Proc. Natl. Acad. Sci. USA 1990, 87, 4828–4832. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current understanding of RANK signaling in osteoclast differentiation and maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [PubMed] [Green Version]
- Lee, K.; Seo, I.; Choi, M.H.; Jeong, D. Roles of mitogen-activated protein kinases in osteoclast biology. Int. J. Mol. Sci. 2018, 19, 3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iotsova, V.; Caamaño, J.; Loy, J.; Yang, Y.; Lewin, A.; Bravo, R. Osteopetrosis in mice lacking NF-kappaB1 and NF-kappaB2. Nat. Med. 1997, 3, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in osteoclast differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Q.; Ovitt, C.; Grigoriadis, A.E.; Mohle-Steinlein, U.; Ruther, U.; Wagner, E.F. Bone and haematopoietic defects in mice lacking c-fos. Nature 1992, 360, 741–745. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Aliprantis, A.O.; Ueki, Y.; Sulyanto, R.; Park, A.; Sigrist, K.S.; Sharma, S.M.; Ostrowski, M.C.; Olsen, B.R.; Glimcher, L.H. NFATc1 in mice represses osteoprotegerin during osteoclastogenesis and dissociates systemic osteopenia from inflammation in cherubism. J. Clin. Investig. 2008, 118, 3775–3789. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; Laplace, C.; Wang, K.Z.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H.; et al. Nuclear factor of activated T-cells (NFAT) rescues osteoclastogenesis in precursors lacking c-Fos. J. Biol. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [Green Version]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Gravallese, E.M.; Harada, Y.; Wang, J.T.; Gorn, A.H.; Thornhill, T.S.; Goldring, S.R. Identification of cell types responsible for bone resorption in rheumatoid arthritis and juvenile rheumatoid arthritis. Am. J. Pathol. 1998, 152, 943–951. [Google Scholar] [PubMed]
- Okada, Y.; Naka, K.; Kawamura, K.; Matsumoto, T.; Nakanishi, I.; Fujimoto, N.; Sato, H.; Seiki, M. Localization of matrix metalloproteinase 9 (92-kilodalton gelatinase/type IV collagenase = gelatinase B) in osteoclasts: Implications for bone resorption. Lab Investig. 1995, 72, 311–322. [Google Scholar] [PubMed]
- Barrow, A.D.; Raynal, N.; Andersen, T.L.; Slatter, D.A.; Bihan, D.; Pugh, N.; Cella, M.; Kim, T.; Rho, J.; Negishi-Koga, T.; et al. OSCAR is a collagen receptor that costimulates osteoclastogenesis in DAP12-deficient humans and mice. J. Clin. Investig. 2011, 121, 3505–3516. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Lee, S.H.; Ha Kim, J.; Choi, Y.; Kim, N. NFATc1 induces osteoclast fusion via up-regulation of Atp6v0d2 and the dendritic cell-specific transmembrane protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Chen, X.C.; Song, J.Y.; Pang, X.H.; Chen, S.L. Internal transcribed spacer 2 barcode: A good tool for identifying Acanthopanacis cortex. Front. Plant Sci. 2015, 6, 840. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Xu, J.; Chen, F.; Liu, T.; Li, J.; Jiang, L.; Jia, Y.; Hu, C.; Gao, Z.; Gan, C.; et al. Acanthopanax senticosus aqueous extract ameliorates ovariectomy-induced bone loss in middle-aged mice by inhibiting the receptor activator of nuclear factor-κB ligand-induced osteoclastogenesis. Food Funct. 2020, 11, 9696–9709. [Google Scholar] [CrossRef] [PubMed]
- Kalu, D.N. The ovariectomized rat model of postmenopausal bone loss. Bone Min. 1991, 15, 175–191. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′-3′) | |
|---|---|---|
| c-Fos | Forward | CTGGTGCAGCCCACTCTGGTC |
| Reverse | CTTTCAGCAGATTGGCAATCTC | |
| NFATc1 | Forward | CAACGCCCTGACCACCGATAG |
| Reverse | GGCTGCCTTCCGTCTCATAGT | |
| TRAP | Forward | ACTTCCCCAGCCCTTACTAC |
| Reverse | TCAGCACATAGCCCACACCG | |
| OSCAR | Forward | CTGCTGGTAACGGATCAGCTCCCCAGA |
| Reverse | CCAAGGAGCCAGAACCTTCGAAACT | |
| Atp6v0d2 | Forward | TCAGATCTCTTCAAGGCTGTGCTG |
| Reverse | GTGCCAAATGAGTTCAGAGTGATG | |
| Cathepsin K | Forward | ACGGAGGCATTGACTCTGAAGATG |
| Reverse | GTTGTTCTTATTCCGAGCCAAGAG | |
| MMP-9 | Forward | TCCAACCTCACGGACACCC |
| Reverse | AGCAAAGCCGGCCGTAGA | |
| Calcitonin receptor | Forward | TCCAACAAGGTGCTTGGGAA |
| Reverse | CTTGAACTGCGTCCACTGGG | |
| DC-STAMP | Forward | TCCTCCATGAACAAACAGTTCCA |
| Reverse | AGACGTGGTTTAGGAATGCAGCTC | |
| OC-STAMP | Forward | ATGAGGACCATCAGGGCAGCCACG |
| Reverse | GGAGAAGCTGGGTCAGTAGTTCGT | |
| GAPDH | Forward | ACCACAGTCCATGCCATCAC |
| Reverse | TCCACCACCCTGTTGCTGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.-Y.; Kim, J.-H.; Jo, E.-H.; Kim, Y.-K. Eleutherococcus sessiliflorus Inhibits Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-Induced Osteoclast Differentiation and Prevents Ovariectomy (OVX)-Induced Bone Loss. Molecules 2021, 26, 1886. https://doi.org/10.3390/molecules26071886
Han S-Y, Kim J-H, Jo E-H, Kim Y-K. Eleutherococcus sessiliflorus Inhibits Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-Induced Osteoclast Differentiation and Prevents Ovariectomy (OVX)-Induced Bone Loss. Molecules. 2021; 26(7):1886. https://doi.org/10.3390/molecules26071886
Chicago/Turabian StyleHan, Sang-Yong, June-Hyun Kim, Eun-Heui Jo, and Yun-Kyung Kim. 2021. "Eleutherococcus sessiliflorus Inhibits Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-Induced Osteoclast Differentiation and Prevents Ovariectomy (OVX)-Induced Bone Loss" Molecules 26, no. 7: 1886. https://doi.org/10.3390/molecules26071886