Cyclin Y Is Expressed in Platelets and Modulates Integrin Outside-in Signaling

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
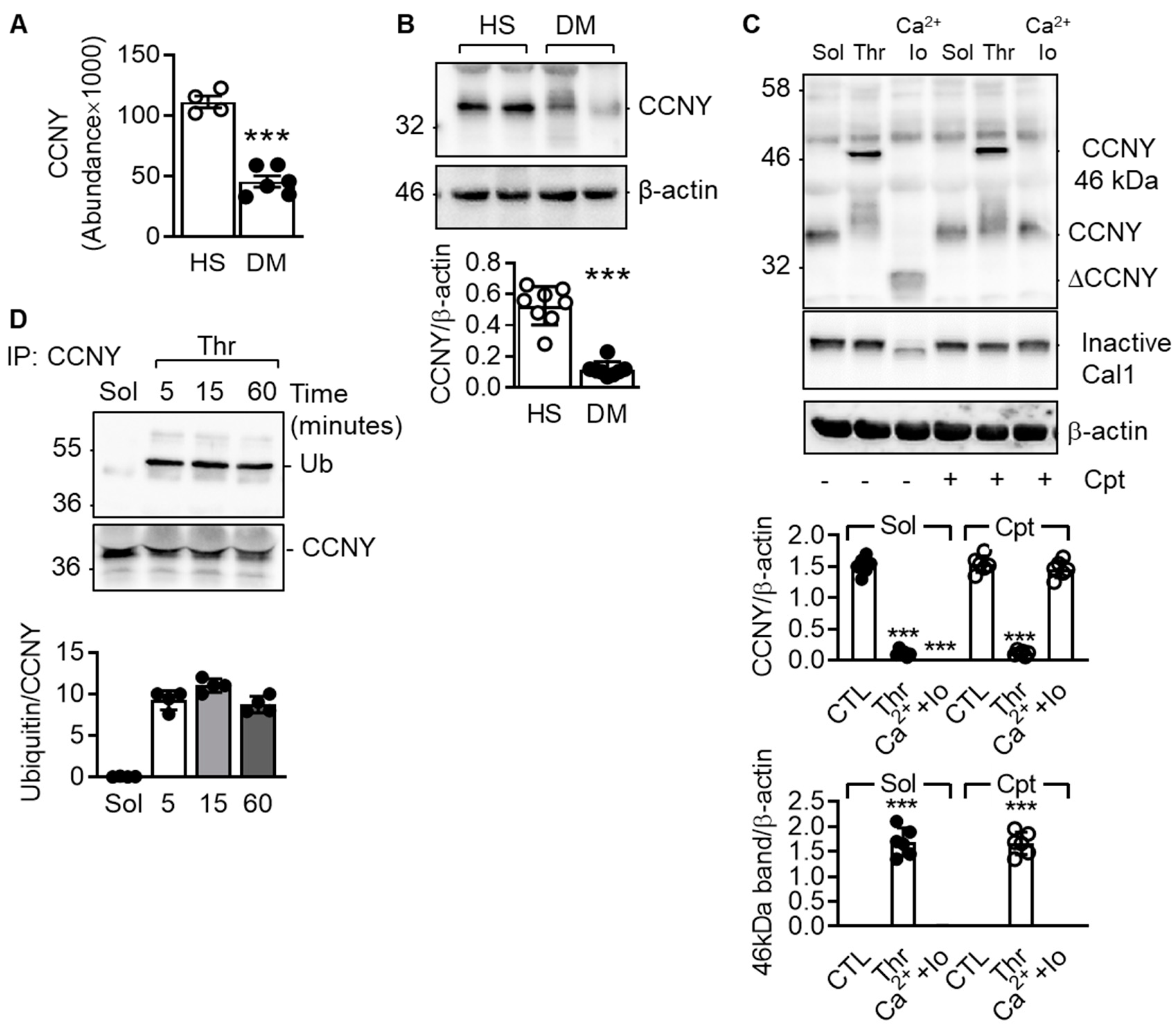
2.1. Cyclin Y Is Expressed in Platelets and Is Ubiquitinated in Response to Platelet Stimulation by Thrombin
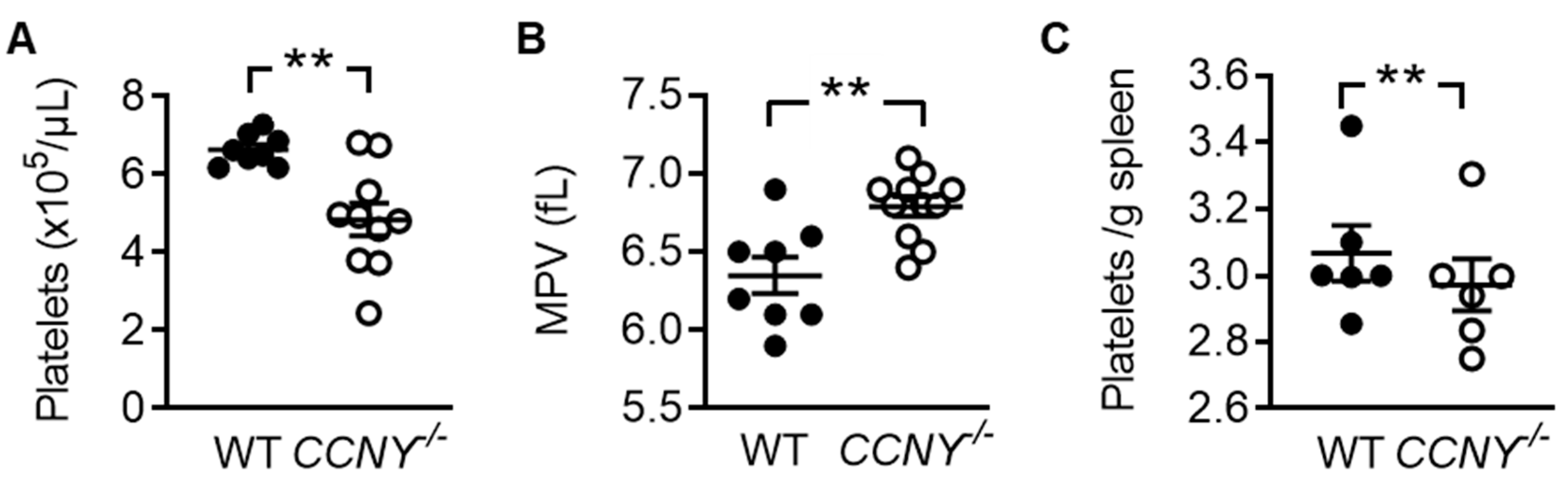
2.2. Cyclin Y Is Involved in Platelet Biogenesis and Function
2.3. CCNY Regulates β3 Integrin Tyrosine Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Study Subjects
4.3. Animals
4.4. Blood Counting
4.5. Flow Cytometry
4.6. Platelet Isolation
4.7. Platelet Aggregation
4.8. ATP Assay
4.9. P-selectin Expression
4.10. Platelet Adhesion and Spreading Assays
4.11. Clot Retraction
4.12. Bleeding Time
4.13. LC-MS-based Proteomics
4.14. Proteomic Data Analysis
4.15. Activation of Calpain In Vitro
4.16. Western Blotting
4.17. Immunoprecipitation
4.18. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Smyth, S.S.; McEver, R.P.; Weyrich, A.S.; Morrell, C.N.; Hoffman, M.R.; Arepally, G.M.; French, P.A.; Dauerman, H.L.; Becker, R.C. Platelet functions beyond hemostasis. J. Thromb. Haemost. 2009, 7, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, M. Role of platelets and platelet receptors in cancer metastasis. J. Hematol. Oncol. 2018, 11, 125. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.W.; Chappaz, S.; Corduan, A.; Chong, M.M.; Campbell, R.; Khoury, A.; Manne, B.K.; Wurtzel, J.G.; Michael, J.V.; Goldfinger, L.E.; et al. Dicer1-mediated miRNA processing shapes the mRNA profile and function of murine platelets. Blood 2016, 127, 1743–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyrich, A.S.; Schwertz, H.; Kraiss, L.W.; Zimmerman, G.A. Protein synthesis by platelets: Historical and new perspectives. J. Thromb. Haemost. 2009, 7, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Denis, M.M.; Tolley, N.D.; Bunting, M.; Schwertz, H.; Jiang, H.; Lindemann, S.; Yost, C.C.; Rubner, F.J.; Albertine, K.H.; Swoboda, K.J.; et al. Escaping the nuclear confines: Signal-dependent pre-mRNA splicing in anucleate platelets. Cell 2005, 122, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Lannan, K.L.; Sahler, J.; Kim, N.; Spinelli, S.L.; Maggirwar, S.B.; Garraud, O.; Cognasse, F.; Blumberg, N.; Phipps, R.P. Breaking the mold: Transcription factors in the anucleate platelet and platelet-derived microparticles. Front. Immunol. 2015, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Schedel, A.; Thornton, S.; Schloss, P.; Kluter, H.; Bugert, P. Human platelets express functional alpha7-nicotinic acetylcholine receptors. Arter. Thromb. Vasc. Biol. 2011, 31, 928–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaurasia, S.N.; Kushwaha, G.; Pandey, A.; Dash, D. Human platelets express functional ectonucleotidases that restrict platelet activation signaling. Biochem. Biophys. Res. Commun. 2020, 527, 104–109. [Google Scholar] [CrossRef]
- Kyselova, A.; Zukunft, S.; Puppe, D.; Wittig, I.; Mann, W.A.; Dornauf, I.; Fleming, I.; Randriamboavonjy, V. Human platelets are a source of collagen I. Haematologica 2020. [Google Scholar] [CrossRef]
- Senzel, L.; Gnatenko, D.V.; Bahou, W.F. The platelet proteome. Curr. Opin. Hematol. 2009, 16, 329–333. [Google Scholar] [CrossRef]
- Loroch, S.; Trabold, K.; Gambaryan, S.; Reiss, C.; Schwierczek, K.; Fleming, I.; Sickmann, A.; Behnisch, W.; Zieger, B.; Zahedi, R.P.; et al. Alterations of the platelet proteome in type I Glanzmann thrombasthenia caused by different homozygous delG frameshift mutations in ITGA2B. Thromb. Haemost. 2017, 117, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Sabrkhany, S.; Kuijpers, M.J.E.; Knol, J.C.; Olde Damink, S.W.M.; Dingemans, A.C.; Verheul, H.M.; Piersma, S.R.; Pham, T.V.; Griffioen, A.W.; Oude Egbrink, M.G.A.; et al. Exploration of the platelet proteome in patients with early-stage cancer. J. Proteom. 2018, 177, 65–74. [Google Scholar] [CrossRef]
- Randriamboavonjy, V.; Fleming, I. Platelet Function and Signaling in Diabetes Mellitus. Curr. Vasc. Pharmacol. 2012. [Google Scholar] [CrossRef]
- Randriamboavonjy, V.; Isaak, J.; Elgheznawy, A.; Pistrosch, F.; Fromel, T.; Yin, X.; Badenhoop, K.; Heide, H.; Mayr, M.; Fleming, I. Calpain inhibition stabilizes the platelet proteome and reactivity in diabetes. Blood 2012, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Randriamboavonjy, V.; Fleming, I. All cut up! The consequences of calpain activation on platelet function. Vascul. Pharmacol. 2012, 56, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A. Mechanisms and regulation of ubiquitin-mediated cyclin degradation. Adv. Exp. Med. Biol. 1996, 389, 221–227. [Google Scholar] [CrossRef]
- Obaya, A.J.; Sedivy, J.M. Regulation of cyclin-Cdk activity in mammalian cells. Cell. Mol. Life Sci. 2002, 59, 126–142. [Google Scholar] [CrossRef]
- Qie, S.; Diehl, J.A. Cyclin D degradation by E3 ligases in cancer progression and treatment. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- An, W.; Zhang, Z.; Zeng, L.; Yang, Y.; Zhu, X.; Wu, J. Cyclin Y Is Involved in the Regulation of Adipogenesis and Lipid Production. PLoS ONE 2015, 10, e0132721. [Google Scholar] [CrossRef]
- Payrastre, B.; Missy, K.; Trumel, C.; Bodin, S.; Plantavid, M.; Chap, H. The integrin alpha IIb/beta 3 in human platelet signal transduction. Biochem. Pharmacol. 2000, 60, 1069–1074. [Google Scholar] [CrossRef]
- Akbiyik, F.; Ray, D.M.; Gettings, K.F.; Blumberg, N.; Francis, C.W.; Phipps, R.P. Human bone marrow megakaryocytes and platelets express PPARgamma, and PPARgamma agonists blunt platelet release of CD40 ligand and thromboxanes. Blood 2004, 104, 1361–1368. [Google Scholar] [CrossRef] [Green Version]
- Gambaryan, S.; Kobsar, A.; Rukoyatkina, N.; Herterich, S.; Geiger, J.; Smolenski, A.; Lohmann, S.M.; Walter, U. Thrombin and collagen induce a feedback inhibitory signaling pathway in platelets involving dissociation of the catalytic subunit of protein kinase A from an NFkappaB-IkappaB complex. J. Biol. Chem. 2010, 285, 18352–18363. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Guest, S.; Finley, R.L., Jr. Why cyclin Y? A highly conserved cyclin with essential functions. Fly (Austin.) 2010, 4, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, Z.; Wang, J.; Li, J.; Wang, H.; Yue, W. Lentivirus-mediated knockdown of cyclin Y (CCNY) inhibits glioma cell proliferation. Oncol. Res. 2010, 18, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Zhao, X.; Zhang, L.; Xu, S.; Liu, Z.; Ma, L.; Jia, W.; Qian, Z.; Zhang, C.; Wang, Y.; et al. Cell cycle protein cyclin Y is associated with human non-small-cell lung cancer proliferation and tumorigenesis. Clin. Lung Cancer 2011, 12, 43–50. [Google Scholar] [CrossRef]
- Liu, H.; Shi, H.; Fan, Q.; Sun, X. Cyclin Y regulates the proliferation, migration, and invasion of ovarian cancer cells via Wnt signaling pathway. Tumour. Biol. 2016, 37, 10161–10175. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Cai, C.; Li, S.; Wang, W.; Li, Y.; Chen, J.; Zhu, X.; Zeng, Y.A. Essential Roles of Cyclin Y-Like 1 and Cyclin Y in Dividing Wnt-Responsive Mammary Stem/Progenitor Cells. PLoS Genet. 2016, 12, e1006055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joe, I.S.; Kim, J.H.; Kim, H.; Hong, J.H.; Kim, M.; Park, M. Cyclin Y-mediated transcript profiling reveals several important functional pathways regulated by Cyclin Y in hippocampal neurons. PLoS ONE 2017, 12, e0172547. [Google Scholar] [CrossRef] [Green Version]
- Ou, C.Y.; Poon, V.Y.; Maeder, C.I.; Watanabe, S.; Lehrman, E.K.; Fu, A.K.; Park, M.; Fu, W.Y.; Jorgensen, E.M.; Ip, N.Y.; et al. Two cyclin-dependent kinase pathways are essential for polarized trafficking of presynaptic components. Cell 2010, 141, 846–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.; Watanabe, S.; Poon, V.Y.; Ou, C.Y.; Jorgensen, E.M.; Shen, K. CYY-1/cyclin Y and CDK-5 differentially regulate synapse elimination and formation for rewiring neural circuits. Neuron 2011, 70, 742–757. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Zimmet, J.M.; Toselli, P.; Thompson, A.; Jackson, C.W.; Ravid, K. Overexpression of cyclin D1 moderately increases ploidy in megakaryocytes. Haematologica 2001, 86, 17–23. [Google Scholar]
- Bennett, J.S. Structure and function of the platelet integrin alphaIIbbeta3. J. Clin. Investig. 2005, 115, 3363–3369. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.L.; Mascelli, M.A.; Neblock, D.S.; Weisman, H.F.; Coller, B.S.; Jordan, R.E. Analysis of GPIIb/IIIa receptor number by quantification of 7E3 binding to human platelets. Blood 1996, 88, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.B.; Smith, D.M.; Plow, E.F. Analysis of Fyn function in hemostasis and alphaIIbbeta3-integrin signaling. J. Cell Sci. 2008, 121, 1641–1648. [Google Scholar] [CrossRef] [Green Version]
- Sadowski, M.; Suryadinata, R.; Tan, A.R.; Roesley, S.N.; Sarcevic, B. Protein monoubiquitination and polyubiquitination generate structural diversity to control distinct biological processes. IUBMB Life 2012, 64, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Jura, N.; Scotto-Lavino, E.; Sobczyk, A.; Bar-Sagi, D. Differential modification of Ras proteins by ubiquitination. Mol. Cell 2006, 21, 679–687. [Google Scholar] [CrossRef]
- Pavri, R.; Zhu, B.; Li, G.; Trojer, P.; Mandal, S.; Shilatifard, A.; Reinberg, D. Histone H2B monoubiquitination functions cooperatively with FACT to regulate elongation by RNA polymerase II. Cell 2006, 125, 703–717. [Google Scholar] [CrossRef] [Green Version]
- Randriamboavonjy, V.; Isaak, J.; Fromel, T.; Viollet, B.; Fisslthaler, B.; Preissner, K.T.; Fleming, I. AMPK alpha2 subunit is involved in platelet signaling, clot retraction, and thrombus stability. Blood 2010, 116, 2134–2140. [Google Scholar] [CrossRef] [Green Version]
- Randriamboavonjy, V.; Schrader, J.; Busse, R.; Fleming, I. Insulin induces the release of vasodilator compounds from platelets by a nitric oxide-G kinase-VAMP-3-dependent pathway. J. Exp. Med. 2004, 199, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Solari, F.A.; Mattheij, N.J.; Burkhart, J.M.; Swieringa, F.; Collins, P.W.; Cosemans, J.M.; Sickmann, A.; Heemskerk, J.W.; Zahedi, R.P. Combined Quantification of the Global Proteome, Phosphoproteome, and Proteolytic Cleavage to Characterize Altered Platelet Functions in the Human Scott Syndrome. Mol. Cell. Proteom. 2016, 15, 3154–3169. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Burkhart, J.M.; Schumbrutzki, C.; Wortelkamp, S.; Sickmann, A.; Zahedi, R.P. Systematic and quantitative comparison of digest efficiency and specificity reveals the impact of trypsin quality on MS-based proteomics. J. Proteom. 2012, 75, 1454–1462. [Google Scholar] [CrossRef]
- Olsen, J.V.; de Godoy, L.M.; Li, G.; Macek, B.; Mortensen, P.; Pesch, R.; Makarov, A.; Lange, O.; Horning, S.; Mann, M. Parts per million mass accuracy on an Orbitrap mass spectrometer via lock mass injection into a C-trap. Mol. Cell. Proteom. 2005, 4, 2010–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaudel, M.; Barsnes, H.; Berven, F.S.; Sickmann, A.; Martens, L. SearchGUI: An open-source graphical user interface for simultaneous OMSSA and X!Tandem searches. Proteomics 2011, 11, 996–999. [Google Scholar] [CrossRef]
- Vaudel, M.; Burkhart, J.M.; Zahedi, R.P.; Oveland, E.; Berven, F.S.; Sickmann, A.; Martens, L.; Barsnes, H. PeptideShaker enables reanalysis of MS-derived proteomics data sets. Nat. Biotechnol. 2015, 33, 22–24. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Fleming, I.; Fisslthaler, B.; Dixit, M.; Busse, R. Role of PECAM-1 in the shear-stress-induced activation of Akt and the endothelial nitric oxide synthase (eNOS) in endothelial cells. J. Cell Sci. 2005, 118, 4103–4111. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyselova, A.; Siragusa, M.; Anthes, J.; Solari, F.A.; Loroch, S.; Zahedi, R.P.; Walter, U.; Fleming, I.; Randriamboavonjy, V. Cyclin Y Is Expressed in Platelets and Modulates Integrin Outside-in Signaling. Int. J. Mol. Sci. 2020, 21, 8239. https://doi.org/10.3390/ijms21218239
Kyselova A, Siragusa M, Anthes J, Solari FA, Loroch S, Zahedi RP, Walter U, Fleming I, Randriamboavonjy V. Cyclin Y Is Expressed in Platelets and Modulates Integrin Outside-in Signaling. International Journal of Molecular Sciences. 2020; 21(21):8239. https://doi.org/10.3390/ijms21218239
Chicago/Turabian StyleKyselova, Anastasia, Mauro Siragusa, Julian Anthes, Fiorella Andrea Solari, Stefan Loroch, René P. Zahedi, Ulrich Walter, Ingrid Fleming, and Voahanginirina Randriamboavonjy. 2020. "Cyclin Y Is Expressed in Platelets and Modulates Integrin Outside-in Signaling" International Journal of Molecular Sciences 21, no. 21: 8239. https://doi.org/10.3390/ijms21218239