
Identification and Characterization of Three New Antimicrobial Peptides from the Marine Mollusk Nerita versicolor (Gmelin, 1791)
 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Peptide Sequencing Analysis with NanoLC-ESI-MS-MS
2.2. Structural Predictions
2.3. Refolding Near a Membrane
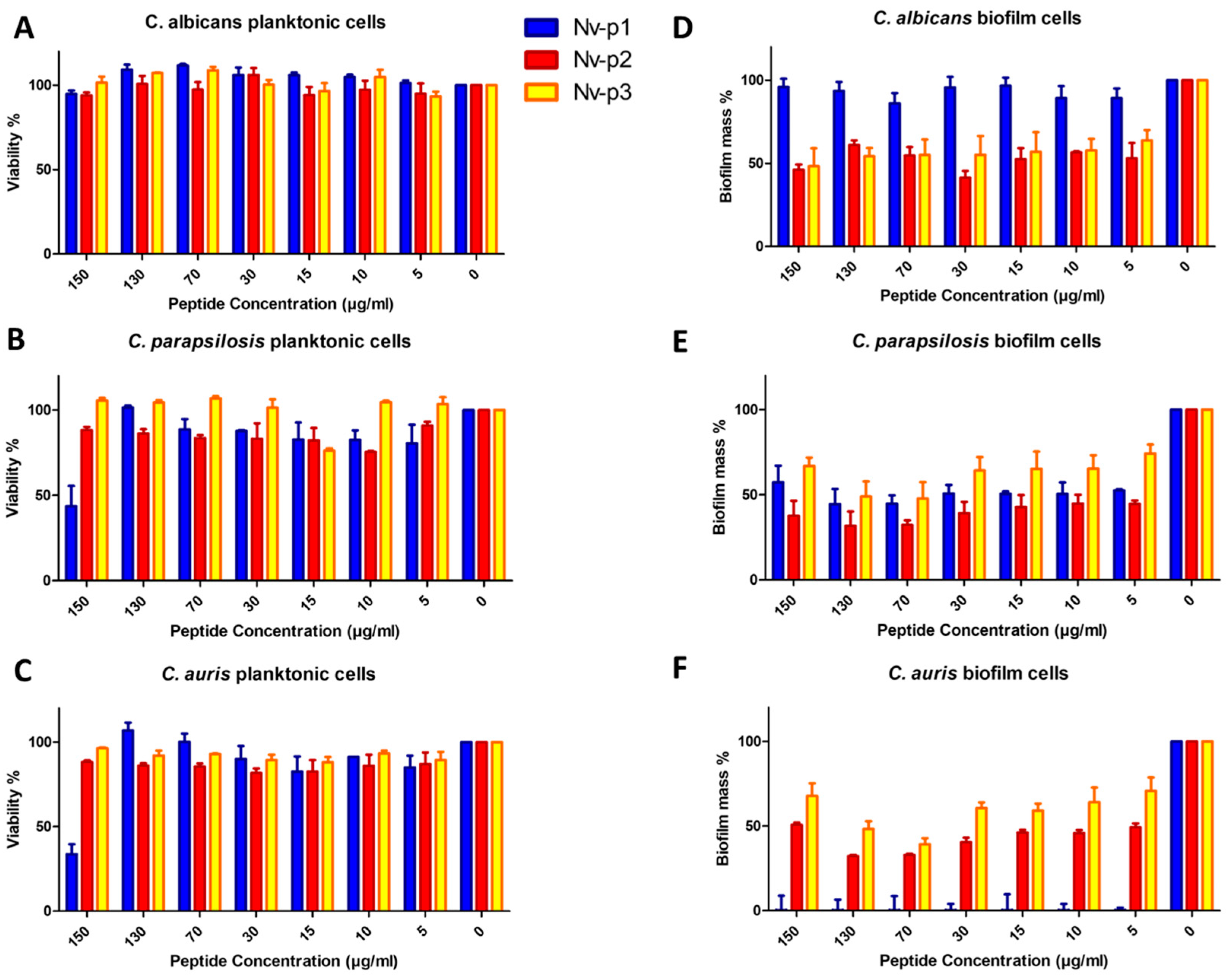
2.4. Antimicrobial Activity
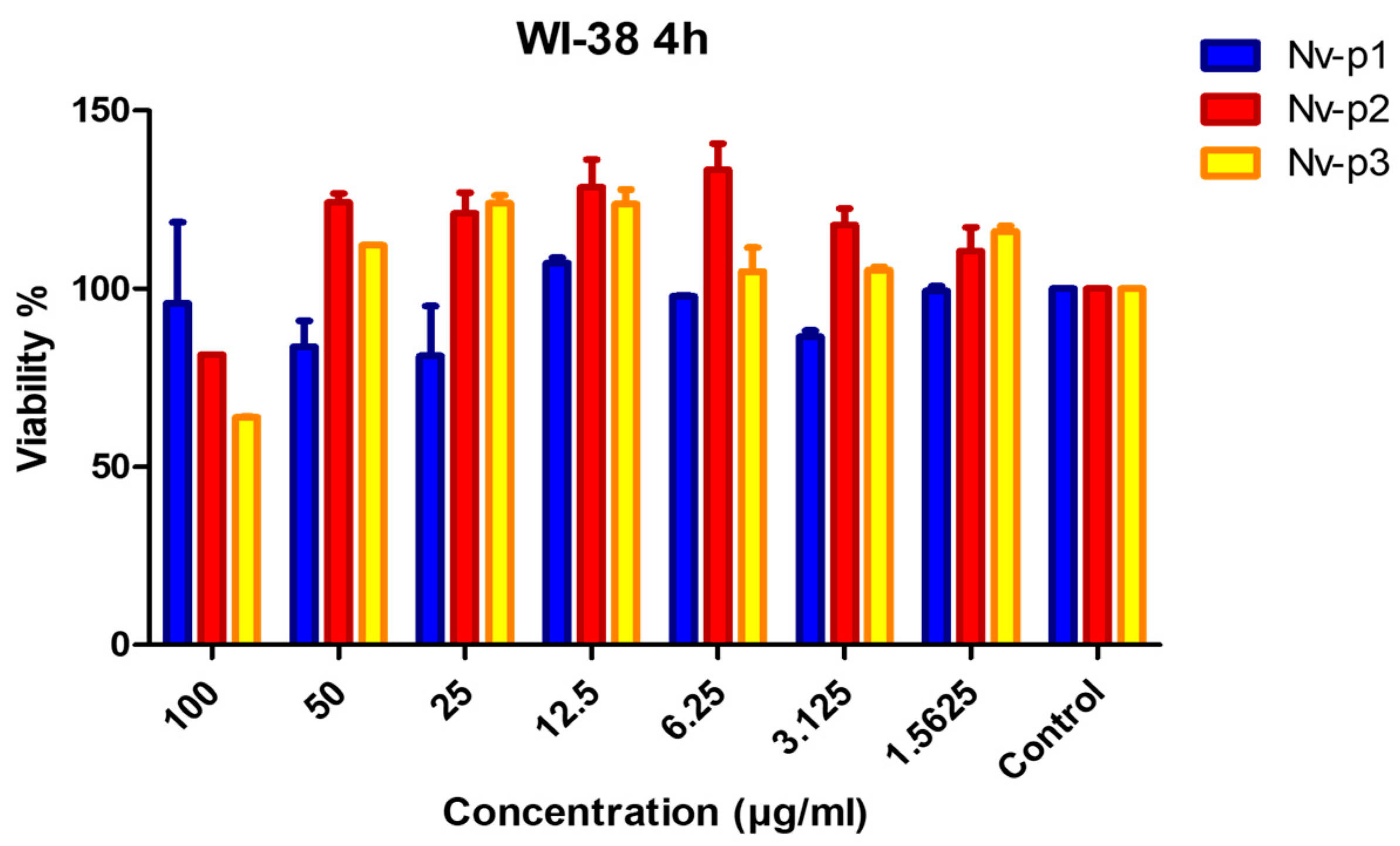
2.5. Toxicity Assays
3. Discussion
4. Materials and Methods
4.1. Invertebrate Collection and Sample Preparation
4.2. Microorganism Strains and Growth Conditions
4.3. Human Cells and Culture Conditions
4.4. Peptide Sequencing Analysis with NanoLC-ESI-MS-MS
4.5. Antimicrobial Activity Prediction
4.6. Peptide Synthesis
4.7. Structural Prediction
4.8. Determination of Antimicrobial Activity
4.9. 3H-Uracil Proliferation Assay
4.10. Antifungal Bioassays
4.11. Biofilm Formation and Quantification/AntiBiofilm Treatment
4.12. Toxicity Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-Membrane Permeabilizing Modes of Action of Antimicrobial Peptides on Bacteria. Curr. Top. Med. Chem. 2016, 16, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.T.; Gellatly, S.L.; Hancock, R.E. Multifunctional cationic host defense peptides and their clinical applications. Cell. Mol. Life Sci. 2011, 68, 2161–2176. [Google Scholar] [CrossRef] [PubMed]
- Omardien, S.; Brul, S.; Zaat, S.A.J. Antimicrobial Activity of Cationic Antimicrobial Peptides against Gram-Positives: Current Progress Made in Understanding the Mode of Action and the Response of Bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Otvos, L., Jr. Immunomodulatory Effects of Antimicrobial Peptides. Acta Microbiol. Immunol. Hung. 2016, 63, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qiu, L.; Zhou, Z.; Song, L. Research progress on the mollusk immunity in China. Dev Comp Immunol 2013, 39, 2–10. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Dorrington, T.; Villamil, L.; Gómez-Chiarri, M. Upregulation in response to infection and antibacterial activity of oyster histone H4. Fish Shellfish. Immunol. 2011, 30, 94–101. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Tincu, J.A.; Taylor, S.W. Antimicrobial Peptides from Marine Invertebrates. Antimicrob. Agents Chemother. 2004, 48, 3645–3654. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dorrington, T.; Gómez-Chiarri, M. The role of histones in the immune responses of aquatic invertebrates. Invertebr. Surviv. J. 2013, 10, 94–101. [Google Scholar]
- Dong, W.; Luo, X.; Sun, Y.; Li, Y.; Wang, C.; Guan, Y.; Shang, D. Binding Properties of DNA and Antimicrobial Peptide Chensinin-1b Containing Lipophilic Alkyl Tails. J. Fluoresc. 2020, 30, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y. The Role of Non-Chromosomal Histones in the Host Defense System. Microbiol. Immunol. 2003, 47, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Birkemo, G.A.; Mantzilas, D.; Lüders, T.; Nes, I.F.; Nissen-Meyer, J. Identification and structural analysis of the antimicrobial domain in hipposin, a 51-mer antimicrobial peptide isolated from Atlantic halibut. Biochim. Biophys. Acta 2004, 1699, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Sathyan, N.; Philip, R.; Chaithanya, E.R.; Kumar, P.R.A. Identification and Molecular Characterization of Molluskin, a Histone-H2A-Derived Antimicrobial Peptide from Molluscs. ISRN Mol. Biol. 2012, 2012, 219656. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.; Trinh, T.T.; Le, H.T.; Tang, S.C.; Hincke, M.; Wellman-Labadie, O.; Ziai, S. Antimicrobial effects of H4-(86-100), histogranin and related compounds—Possible involvement of DNA gyrase. FEBS J. 2008, 5, 5286–5297. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Huang, C.-M.; Nakatsuji, T.; Thiboutot, D.; Kang, S.-A.; Monestier, M.; Gallo, R.L. Histone H4 Is a Major Component of the Antimicrobial Action of Human Sebocytes. J. Investig. Dermatol. 2009, 129, 2489–2496. [Google Scholar] [CrossRef]
- Kawasaki, H.; Iwamuro, S. Potential roles of histones in host defense as antimicrobial agents. Infect. Disord. Drug Targets 2008, 3, 195–205. [Google Scholar] [CrossRef]
- Ho, Y.H.; Shah, P.; Chen, Y.W.; Chen, C.S. Systematic Analysis of Intracellular-targeting Antimicrobial Peptides, Bactenecin 7, Hybrid of Pleurocidin and Dermaseptin, Proline-Arginine-rich Peptide, and Lactoferricin B, by Using Escherichia coli Proteome Microarrays. Mol. Cell. Proteom. 2016, 15, 1837–1847. [Google Scholar] [CrossRef]
- Yacoub, T.; Rima, M.; Karam, M.; Sabatier, J.-M.; Fajloun, J.-M.S.A.Z. Antimicrobials from Venomous Animals: An Overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- Samy, R.P.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef]
- Perumal Samy, R.; Pachiappan, A.; Gopalakrishnakone, P.; Thwin, M.M.; Hian, Y.E.; Chow, V.T.K.; Bow, H.; Weng, J.T. In vitro antimicrobial activity of natural toxins and animal venoms tested against Burkholderia pseudomallei. BMC Infect. Dis. 2006, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.; Sali, A. Protein Structure Prediction and Structural Genomics. Science 2001, 294, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Cebrian, R.; Rodriguez-Cabezas, M.E.; Martín-Escolano, R.; Rubiño, S.; Barros, M.G.; Montalbán-López, M.; Rosales, M.J.; Sánchez-Moreno, M.; Valdivia, E.; Martínez-Bueno, M.; et al. Preclinical studies of toxicity and safety of the AS-48 bacteriocin. J. Adv. Res. 2019, 20, 129–139. [Google Scholar] [CrossRef]
- Starr, C.G.; Ghimire, J.; Guha, S.; Hoffmann, J.P.; Wang, Y.; Sun, L.; Landreneau, B.N.; Kolansky, Z.D.; Kilanowski-Doroh, I.M.; Sammarco, M.C.; et al. Synthetic molecular evolution of host cell-compatible, antimicrobial peptides effective against drug-resistant, biofilm-forming bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 8437–8448. [Google Scholar] [CrossRef]
- Groß, R.; Bauer, R.; Krüger, F.; Rücker-Braun, E.; Olari, L.-R.; Ständker, L.; Preising, N.; Rodríguez, A.A.; Conzelmann, C.; Gerbl, F.; et al. A Placenta Derived C-Terminal Fragment of β-Hemoglobin with Combined Antibacterial and Antiviral Activity. Front. Microbiol. 2020, 11, 508. [Google Scholar] [CrossRef]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMP R3: A database on sequences, structures and signatures of antimicrobial peptides: Table 1. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef]
- Veltri, D.; Kamath, U.; Shehu, A. Deep learning improves antimicrobial peptide recognition. Bioinformatics 2018, 34, 2740–2747. [Google Scholar] [CrossRef]
- Meher, P.K.; Sahu, T.K.; Saini, V.; Rao, A.R. Predicting antimicrobial peptides with improved accuracy by incorporating the compositional, physico-chemical and structural features into Chou’s general PseAAC. Sci. Rep. 2017, 7, srep42362. [Google Scholar] [CrossRef]
- González-García, M.; Rodríguez, A.; Alba, A.; Vázquez, A.; Morales-Vicente, F.; Pérez-Erviti, J.; Spellerberg, B.; Stenger, S.; Grieshober, M.; Conzelmann, C.; et al. New Antibacterial Peptides from the Freshwater Mollusk Pomacea poeyana (Pilsbry, 1927). Biomolecules 2020, 10, 1473. [Google Scholar] [CrossRef]
- Jose, A.; Coco, B.J.; Milligan, S.; Young, B.; Lappin, D.F.; Bagg, J.; Murray, C.; Ramage, G. Reducing the Incidence of Denture Stomatitis: Are Denture Cleansers Sufficient? J. Prosthodont. 2010, 19, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm Formation as Microbial Development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Negri, M.; Gonçalves, V.; Silva, S.; Henriques, M.; Azeredo, J.; Oliveira, R. Crystal violet staining to quantify Candida adhesion to epithelial cells. Br. J. Biomed. Sci. 2010, 67, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms formed by Candida albicans bloodstream isolates display phenotypic and transcriptional heterogeneity that are associated with resistance and pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Primary Structure | pI | Charge at pH 7.4 | Molecular Weight |
|---|---|---|---|---|
| Nv-p1 | SGRGKGGKGLGKGGAKRHR | 12.57 | +6.25 | 1864.127 Da |
| Nv-p2 | SGRGKGGKGLGKGGAKRH | 12.51 | +5.44 | 1707.941 Da |
| Nv-p3 | KKKPTKK | 10.57 | +4.141 | 857.0973 Da |
| Peptide Bacterial species | Nv-p1 | Nv-p2 | Nv-p3 |
|---|---|---|---|
| Pseudomonas aeruginosa | >100 µg/mL | >100 µg/mL | >100 µg/mL |
| Peptides | Antifungal Activity MIC (µg/mL) | Antibiofilm Activity IC50 (µg/mL) | ||||
|---|---|---|---|---|---|---|
| C.albicans | C.parapsilosis | C.auris | C.albicans | C.parapsilosis | C.auris | |
| Nv-p1 | ˃100 | ˃100 | ˃100 | ˃100 | 31.9 | 1.8 |
| Nv-p2 | ˃100 | ˃100 | ˃100 | 40.0 | 12.4 | 15.4 |
| Nv-p3 | ˃100 | ˃100 | ˃100 | 57.5 | 84.4 | 57.5 |
| Peptide | Viability (%) |
|---|---|
| Unstimulated | 100 |
| Nv-p1 | 100 |
| Nv-p2 | 100 |
| Nv-p3 | 86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, A.; Martell-Huguet, E.M.; González-García, M.; Alpízar-Pedraza, D.; Alba, A.; Vazquez, A.A.; Grieshober, M.; Spellerberg, B.; Stenger, S.; Münch, J.; et al. Identification and Characterization of Three New Antimicrobial Peptides from the Marine Mollusk Nerita versicolor (Gmelin, 1791). Int. J. Mol. Sci. 2023, 24, 3852. https://doi.org/10.3390/ijms24043852
Rodriguez A, Martell-Huguet EM, González-García M, Alpízar-Pedraza D, Alba A, Vazquez AA, Grieshober M, Spellerberg B, Stenger S, Münch J, et al. Identification and Characterization of Three New Antimicrobial Peptides from the Marine Mollusk Nerita versicolor (Gmelin, 1791). International Journal of Molecular Sciences. 2023; 24(4):3852. https://doi.org/10.3390/ijms24043852
Chicago/Turabian StyleRodriguez, Armando, Ernesto M. Martell-Huguet, Melaine González-García, Daniel Alpízar-Pedraza, Annia Alba, Antonio A. Vazquez, Mark Grieshober, Barbara Spellerberg, Steffen Stenger, Jan Münch, and et al. 2023. "Identification and Characterization of Three New Antimicrobial Peptides from the Marine Mollusk Nerita versicolor (Gmelin, 1791)" International Journal of Molecular Sciences 24, no. 4: 3852. https://doi.org/10.3390/ijms24043852