
Deep Learning-Based Precision Analysis for Acrosome Reaction by Modification of Plasma Membrane in Boar Sperm
 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
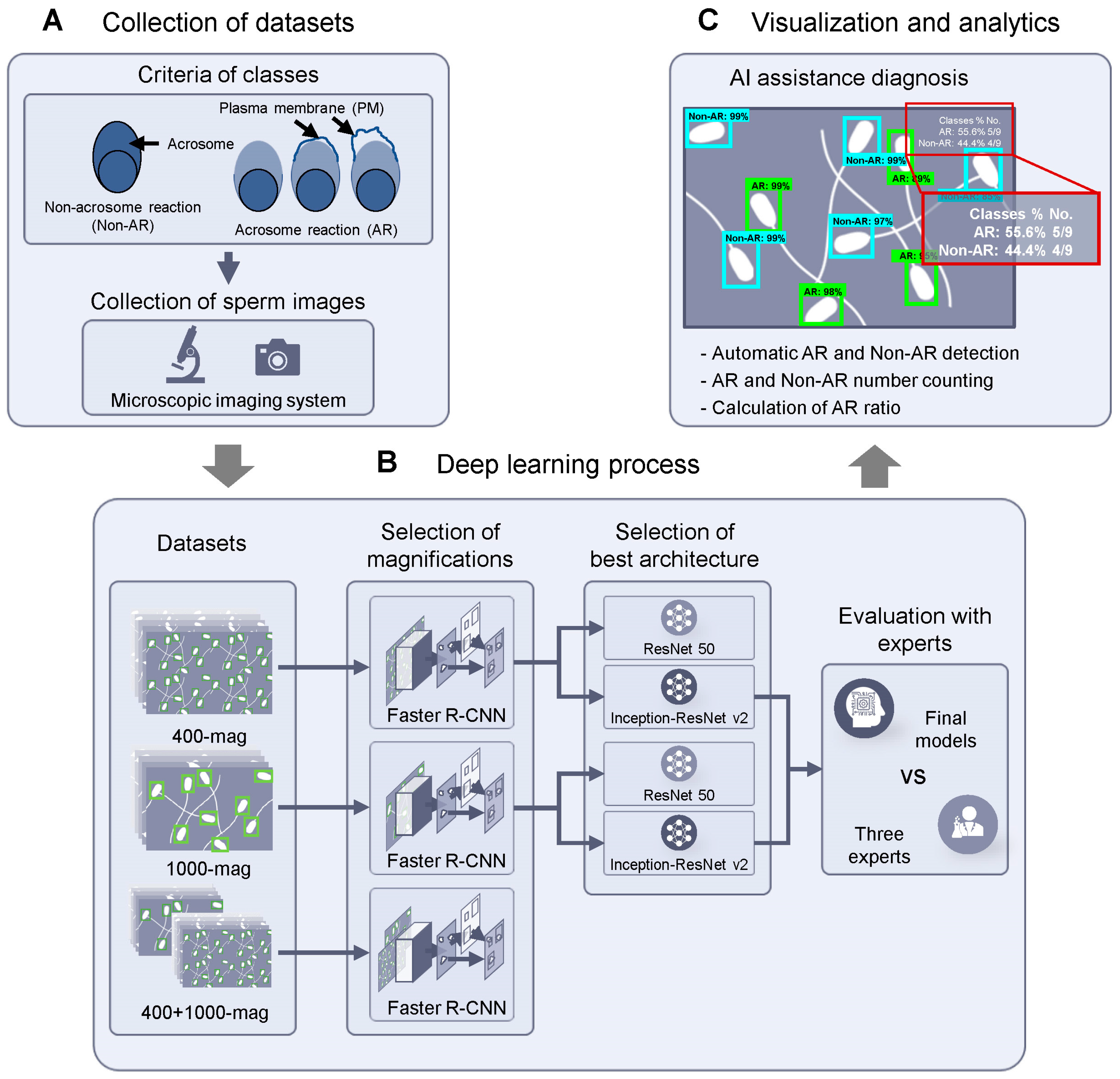
2. Materials and Methods
2.1. Experimental Design
2.2. Sperm Preparation and Dataset Collection
2.3. Data Preparation
2.4. Model Training Using Convolutional Neural Networks (CNNs)
2.5. Comparison of Model Performance with Experts
2.6. Automatic Calculation of Acrosome Reaction Rate
2.7. Statistical Analysis
3. Results
3.1. Selection of Datasets According to Magnifications
3.2. Selection of the Best Architecture
3.3. Comparison of Model Performances with Expert
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tulsiani, D.R.; Abou-Haila, A.; Loeser, C.R.; Pereira, B.M. The biological and functional significance of the sperm acrosome and acrosomal enzymes in mammalian fertilization. Exp. Cell Res. 1998, 240, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Abou-Haila, A.; Tulsiani, D.R. Mammalian sperm acrosome: Formation, contents, and function. Arch. Biochem. Biophys. 2000, 379, 173–182. [Google Scholar] [CrossRef]
- Flesch, F.M.; Gadella, B.M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim. Biophys. Acta Rev. Biomembr. 2000, 1469, 197–235. [Google Scholar] [CrossRef]
- Silva, P.; Gadella, B. Detection of damage in mammalian sperm cells. Theriogenology 2006, 65, 958–978. [Google Scholar] [CrossRef]
- Lee, S.-H.; Park, C.-K. Effect of magnetized extender on sperm membrane integrity and development of oocytes in vitro fertilized with liquid storage boar semen. Anim. Reprod. Sci. 2015, 154, 86–94. [Google Scholar] [CrossRef]
- Vicente-Fiel, S.; Palacin, I.; Santolaria, P.; Yániz, J. A comparative study of sperm morphometric subpopulations in cattle, goat, sheep and pigs using a computer-assisted fluorescence method (CASMA-F). Anim. Reprod. Sci. 2013, 139, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Yániz, J.; Vicente-Fiel, S.; Capistrós, S.; Palacín, I.; Santolaria, P. Automatic evaluation of ram sperm morphometry. Theriogenology 2012, 77, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Ghasemian, F.; Mirroshandel, S.A.; Monji-Azad, S.; Azarnia, M.; Zahiri, Z. An efficient method for automatic morphological abnormality detection from human sperm images. Comput. Methods Programs Biomed. 2015, 122, 409–420. [Google Scholar] [CrossRef]
- Li, J.; Tseng, K.-K.; Dong, H.; Li, Y.; Zhao, M.; Ding, M. Human sperm health diagnosis with principal component analysis and K-nearest neighbor algorithm. In Proceedings of the 2014 International Conference on Medical Biometrics, Shenzhen, China, 30 May–1 June 2014; pp. 108–113. [Google Scholar]
- Shaker, F.; Monadjemi, S.A.; Naghsh-Nilchi, A.R. Automatic detection and segmentation of sperm head, acrosome and nucleus in microscopic images of human semen smears. Comput. Methods Programs Biomed. 2016, 132, 11–20. [Google Scholar] [CrossRef]
- Zhang, Y. Animal sperm morphology analysis system based on computer vision. In Proceedings of the 2017 Eighth International Conference on Intelligent Control and Information Processing (ICICIP), Hangzhou, China, 3–5 November 2017; pp. 338–341. [Google Scholar]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. Imagenet classification with deep convolutional neural networks. In Proceedings of the Advances in Neural Information Processing Systems, Lake Tahoe, NV, USA, 3–6 December 2012; pp. 1097–1105. [Google Scholar]
- Ren, S.; He, K.; Girshick, R.; Sun, J. Faster r-cnn: Towards real-time object detection with region proposal networks. In Proceedings of the Advances in Neural Information Processing Systems, Montreal, QC, Canada, 7–12 December 2015; pp. 91–99. [Google Scholar]
- Simonyan, K.; Zisserman, A. Very deep convolutional networks for large-scale image recognition. arXiv 2014, arXiv:1409.1556. [Google Scholar]
- Girshick, R. Fast r-cnn. In Proceedings of the IEEE International Conference on Computer Vision, Santiago, Chile, 11–18 December 2015; pp. 1440–1448. [Google Scholar]
- Dai, J.; Li, Y.; He, K.; Sun, J. R-fcn: Object detection via region-based fully convolutional networks. In Proceedings of the Advances in Neural Information Processing Systems, Barcelona, Spain, 5–10 December 2016; pp. 379–387. [Google Scholar]
- Redmon, J.; Divvala, S.; Girshick, R.; Farhadi, A. You only look once: Unified, real-time object detection. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 779–788. [Google Scholar]
- Liu, W.; Anguelov, D.; Erhan, D.; Szegedy, C.; Reed, S.; Fu, C.-Y.; Berg, A.C. Ssd: Single shot multibox detector. In Proceedings of the European Conference on Computer Vision, Amsterdam, The Netherlands, 11–12 October 2016; pp. 21–37. [Google Scholar]
- Lin, T.-Y.; Goyal, P.; Girshick, R.; He, K.; Dollár, P. Focal loss for dense object detection. In Proceedings of the IEEE International Conference on Computer Vision, Venice, Italy, 22–29 October 2017; pp. 2980–2988. [Google Scholar]
- Everingham, M.; Eslami, S.A.; Van Gool, L.; Williams, C.K.; Winn, J.; Zisserman, A. The pascal visual object classes challenge: A retrospective. Int. J. Comput. Vis. 2015, 111, 98–136. [Google Scholar] [CrossRef]
- Lee, C.; Kim, H.J.; Oh, K.W. Comparison of faster R-CNN models for object detection. In Proceedings of the 2016 16th International Conference on Control, Automation and Systems (ICCAS), Gyeongju, Republic of Korea, 16–19 October 2016; pp. 107–110. [Google Scholar]
- Wang, Q.; Bi, S.; Sun, M.; Wang, Y.; Wang, D.; Yang, S. Deep learning approach to peripheral leukocyte recognition. PLoS ONE 2019, 14, e0218808. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zeng, X.; Sigmund, S.E.; Lin, R.; Zhou, B.; Liu, C.; Wang, K.; Jiang, R.; Freyberg, Z.; Lv, H. Automatic localization and identification of mitochondria in cellular electron cryo-tomography using faster-RCNN. BMC Bioinform. 2019, 20, 132. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Q. Detection and Classification of Cervical Exfoliated Cells Based on Faster R-CNN. In Proceedings of the 2019 IEEE 11th International Conference on Advanced Infocomm Technology (ICAIT), Jinan, China, 18–19 October 2019; pp. 52–57. [Google Scholar]
- Bianco, S.; Cadene, R.; Celona, L.; Napoletano, P. Benchmark analysis of representative deep neural network architectures. IEEE Access 2018, 6, 64270–64277. [Google Scholar] [CrossRef]
- Szegedy, C.; Liu, W.; Jia, Y.; Sermanet, P.; Reed, S.; Anguelov, D.; Erhan, D.; Vanhoucke, V.; Rabinovich, A. Going deeper with convolutions. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Boston, MA, USA, 7–12 June 2015; pp. 1–9. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Szegedy, C.; Ioffe, S.; Vanhoucke, V.; Alemi, A.A. Inception-ResNet and the Impact of Residual Connections on Learning. In Proceedings of the Thirty-First AAAI Conference on Artificial Intelligence, San Francisco, CA, USA, 4–9 February 2017. [Google Scholar]
- Moen, E.; Bannon, D.; Kudo, T.; Graf, W.; Covert, M.; Van Valen, D. Deep learning for cellular image analysis. Nat. Methods 2019, 16, 1233–1246. [Google Scholar] [CrossRef]
- McCallum, C.; Riordon, J.; Wang, Y.; Kong, T.; You, J.B.; Sanner, S.; Lagunov, A.; Hannam, T.G.; Jarvi, K.; Sinton, D. Deep learning-based selection of human sperm with high DNA integrity. Commun. Biol. 2019, 2, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javadi, S.; Mirroshandel, S.A. A novel deep learning method for automatic assessment of human sperm images. Comput. Biol. Med. 2019, 109, 182–194. [Google Scholar] [CrossRef]
- Hidayatullah, P.; Wang, X.; Yamasaki, T.; Mengko, T.L.; Munir, R.; Barlian, A.; Sukmawati, E.; Supraptono, S. DeepSperm: A robust and real-time bull sperm-cell detection in densely populated semen videos. arXiv 2020, arXiv:2003.01395. [Google Scholar] [CrossRef]
- Rahimzadeh, M.; Attar, A. Sperm detection and tracking in phase-contrast microscopy image sequences using deep learning and modified CSR-DCF. arXiv 2020, arXiv:2002.04034. [Google Scholar]
- Huang, C.; Li, Y.; Loy, C.C.; Tang, X. Learning Deep Representation for Imbalanced Classification. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 5375–5384. [Google Scholar]
- Wang, Y.-X.; Ramanan, D.; Hebert, M. Learning to model the tail. In Proceedings of the Advances in Neural Information Processing Systems, Long Beach, CA, USA, 4–9 December 2017; pp. 7029–7039. [Google Scholar]
- Yoon, H.; Lee, S.-H.; Park, M. TensorFlow with user friendly Graphical Framework for object detection API. arXiv 2020, arXiv:2006.06385. [Google Scholar]
- Lin, T.-Y.; Maire, M.; Belongie, S.; Hays, J.; Perona, P.; Ramanan, D.; Dollár, P.; Zitnick, C.L. Microsoft coco: Common objects in context. In Proceedings of the European Conference on Computer Vision, Zurich, Switzerland, 6–12 September 2014; pp. 740–755. [Google Scholar]
- Everingham, M.; Van Gool, L.; Williams, C.K.; Winn, J.; Zisserman, A. The pascal visual object classes (voc) challenge. Int. J. Comput. Vis. 2010, 88, 303–338. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.K.; Pasqualotto, F.F.; Nelson, D.R.; Agarwal, A. Relationship between seminal white blood cell counts and oxidative stress in men treated at an infertility clinic. J. Androl. 2001, 22, 575–583. [Google Scholar] [PubMed]
- Du Plessis, S.S.; Gokul, S.; Agarwal, A. Semen hyperviscosity: Causes, consequences, and cures. Front. Biosci. (Elite Ed) 2013, 5, 224–231. [Google Scholar] [PubMed]
- Lafuente, R.; García-Blàquez, N.; Jacquemin, B.; Checa, M.A. Outdoor air pollution and sperm quality. Fertil. Steril. 2016, 106, 880–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Animal ID | No. of 400-mag Dataset | No. of 1000-mag Dataset | ||||
|---|---|---|---|---|---|---|
| Images | AR Sperm | Non-AR Sperm | Images | AR Sperm | Non-AR Sperm | |
| 1 | 21 | 312 | 164 | 45 | 291 | 99 |
| 2 | 22 | 412 | 176 | 42 | 243 | 87 |
| 3 | 20 | 173 | 137 | 45 | 291 | 93 |
| 4 | 23 | 210 | 130 | 45 | 213 | 78 |
| 5 | 22 | 342 | 243 | 45 | 234 | 117 |
| 6 | 21 | 306 | 178 | 42 | 300 | 102 |
| 7 | 22 | 286 | 138 | 45 | 279 | 93 |
| 8 | 21 | 214 | 220 | 42 | 243 | 114 |
| 9 | 21 | 175 | 119 | 45 | 204 | 129 |
| 10 | 22 | 302 | 236 | 42 | 87 | 84 |
| Total | 215 | 2732 | 1741 | 438 | 2385 | 996 |
| Dataset | Folds | Images | No. of Labeling Data | ||
|---|---|---|---|---|---|
| AR | Non-AR | ||||
| 400-mag | Training | Fold 1 | 34 | 421 | 273 |
| Fold 2 | 34 | 442 | 280 | ||
| Fold 3 | 34 | 418 | 274 | ||
| Fold 4 | 35 | 444 | 296 | ||
| Fold 5 | 35 | 469 | 279 | ||
| Test | - | 43 | 538 | 339 | |
| 1000-mag | Training | Fold 1 | 72 | 363 | 183 |
| Fold 2 | 69 | 387 | 153 | ||
| Fold 3 | 69 | 369 | 159 | ||
| Fold 4 | 69 | 399 | 141 | ||
| Fold 5 | 72 | 396 | 168 | ||
| Test | - | 87 | 471 | 192 | |
| 400 + 1000-mag | Training | Fold 1 | 106 | 784 | 456 |
| Fold 2 | 103 | 829 | 433 | ||
| Fold 3 | 103 | 787 | 433 | ||
| Fold 4 | 104 | 843 | 437 | ||
| Fold 5 | 107 | 865 | 447 | ||
| Test | - | 130 | 1009 | 531 | |
| Classes | Model Names | Accuracy | Precision | Recall | F1 | AP |
|---|---|---|---|---|---|---|
| AR | ResNet-400-mag | 0.899 ± 0.028 | 0.939 ± 0.022 | 0.893 ± 0.030 | 0.915 ± 0.020 | 0.954 ± 0.010 |
| ResNet-1000-mag | 0.937 ± 0.016 | 0.932 ± 0.011 | 0.932 ± 0.018 | 0.932 ± 0.012 | 0.968 ± 0.014 | |
| ResNet-400 + 1000-mag | 0.894 ± 0.027 | 0.941 ± 0.013 | 0.888 ± 0.027 | 0.913 ± 0.013 | 0.943 ± 0.014 | |
| Non-AR | ResNet-400-mag | 0.870 ± 0.035 | 0.926 ± 0.015 | 0.861 ± 0.037 | 0.892 ± 0.022 | 0.952 ± 0.012 |
| ResNet-1000-mag | 0.899 ± 0.054 | 0.919 ± 0.042 | 0.893 ± 0.054 | 0.904 ± 0.019 | 0.964 ± 0.021 | |
| ResNet-400 + 1000-mag | 0.857 ± 0.023 | 0.930 ± 0.010 | 0.848 ± 0.023 | 0.887 ± 0.012 | 0.957 ± 0.004 | |
| Average | ResNet-400-mag | 0.884 ± 0.021 | 0.932 ± 0.011 | 0.877 ± 0.023 | 0.903 ± 0.017 | 0.953 ± 0.008 |
| ResNet-1000-mag | 0.918 ± 0.032 | 0.926 ± 0.023 | 0.912 ± 0.033 | 0.918 ± 0.015 | 0.966 ± 0.015 | |
| ResNet-400 + 1000-mag | 0.875 ± 0.013 | 0.935 ± 0.009 | 0.868 ± 0.013 | 0.900 ± 0.008 | 0.950 ± 0.021 |
| Models | Accuracy | Precision | Recall | F1 | mAP |
|---|---|---|---|---|---|
| ResNet-400-mag | 0.884 ± 0.021 | 0.932 ± 0.011 | 0.877 ± 0.023 | 0.903 ± 0.017 | 0.953 ± 0.008 |
| Incep-Res-400-mag | 0.896 ± 0.022 | 0.945 ± 0.016 | 0.891 ± 0.024 | 0.917 ± 0.018 | 0.976 ± 0.007 |
| ResNet-1000-mag | 0.918 ± 0.032 | 0.926 ± 0.023 | 0.912 ± 0.033 | 0.918 ± 0.015 | 0.942 ± 0.026 |
| Incep-Res-1000-mag | 0.919 ± 0.018 | 0.912 ± 0.024 | 0.913 ± 0.018 | 0.912 ± 0.006 | 0.974 ± 0.005 |
| Models | Accuracy | Precision | Recall | F1 | mAP | FPS |
|---|---|---|---|---|---|---|
| ResNet-400-mag | 0.900 | 0.935 | 0.893 | 0.914 | 0.963 | 1.572 ± 0.132 |
| Incep-Res-400-mag | 0.916 | 0.951 | 0.912 | 0.931 | 0.982 | 0.872 ± 0.076 |
| ResNet-1000-mag | 0.927 | 0.911 | 0.920 | 0.915 | 0.965 | 0.450 ± 0.023 |
| Incep-Res-1000-mag | 0.933 | 0.922 | 0.927 | 0.919 | 0.971 | 0.266 ± 0.017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.; Yoon, H.; Kang, B.H.; Lee, H.; An, J.; Lee, T.; Cheong, H.-T.; Lee, S.-H. Deep Learning-Based Precision Analysis for Acrosome Reaction by Modification of Plasma Membrane in Boar Sperm. Animals 2023, 13, 2622. https://doi.org/10.3390/ani13162622
Park M, Yoon H, Kang BH, Lee H, An J, Lee T, Cheong H-T, Lee S-H. Deep Learning-Based Precision Analysis for Acrosome Reaction by Modification of Plasma Membrane in Boar Sperm. Animals. 2023; 13(16):2622. https://doi.org/10.3390/ani13162622
Chicago/Turabian StylePark, Mira, Heemoon Yoon, Byeong Ho Kang, Hayoung Lee, Jisoon An, Taehyun Lee, Hee-Tae Cheong, and Sang-Hee Lee. 2023. "Deep Learning-Based Precision Analysis for Acrosome Reaction by Modification of Plasma Membrane in Boar Sperm" Animals 13, no. 16: 2622. https://doi.org/10.3390/ani13162622