Histon-Acetylierung
Eukaryotische Zellen verpacken ihre DNA, indem sie sie fest um Proteine, die Histone genannt werden, wickeln. Histone sind eine Familie von kleinen, positiv geladenen Proteinen. Die menschlichen Histone sind H1, H2A, H2B, H3 und H4. Da die DNA aufgrund des Phosphat-Zucker-Rückgrats negativ geladen ist, bindet sie sich fest an positiv geladene Histone. Der dabei entstehende DNA-Protein-Komplex wird als Chromatin bezeichnet. Die Grundeinheit des Chromatins ist ein Nukleosom, das aus neun Histonproteinen und 166 Basenpaaren der DNA besteht.
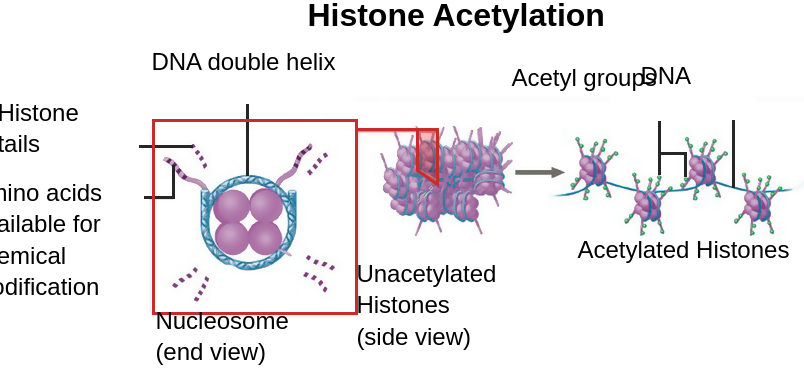
Die Struktur des Chromatins dient nicht nur als Verpackungsmechanismus, sondern auch als Regulator der Genexpression. Die Position der DNA kann die Zugänglichkeit des DNA-Transkriptionsapparats beeinflussen. Wenn sich die DNA in stark kondensiertem Chromatin (Heterochromatin) befindet, ist sie für den DNA-Transkriptionsapparat, wie Transkriptionsfaktoren und die RNA-Polymerase, physisch unzugänglich.
Jedes Histon hat einen 20 Aminosäuren langen „Schwanz“ an seinem N-Terminus, der nach außen ragt. Diese Histonschwänze sind für verschiedene modifizierende Enzyme zugänglich, die zusätzliche spezifische chemische Gruppen wie Acetyl- (-COCH3), Methyl- und Phosphatgruppen entfernen oder katalysieren. Die Histon-Acetyltransferase kann Acetylgruppen an den positiv geladenen Histonschwanz anhängen und damit die Ladung verändern. Durch die Verringerung der positiven Ladung eines Histonschwanzes nimmt die Affinität der DNA zu den Histonen ab, wodurch das verdichtete Chromatin zugänglicher und die Genexpression gefördert wird. Ein anderes Enzym, die Histondeacetylase, bewirkt das Gegenteil der Histon-Acetyltransferase: Dieses Enzym entfernt die Acetylgruppe von den Histonschwänzen und unterdrückt damit die Transkription.
Die Histonacetylierung ist Teil der Genregulation