
Description of the Soil Diatom Sellaphora terrestris sp. nov. (Bacillariophyceae, Sellaphoraceae) from Vietnam, with Remarks on the Phylogeny and Taxonomy of Sellaphora and Systematic Position of Microcostatus

Abstract
:1. Introduction
2. Results
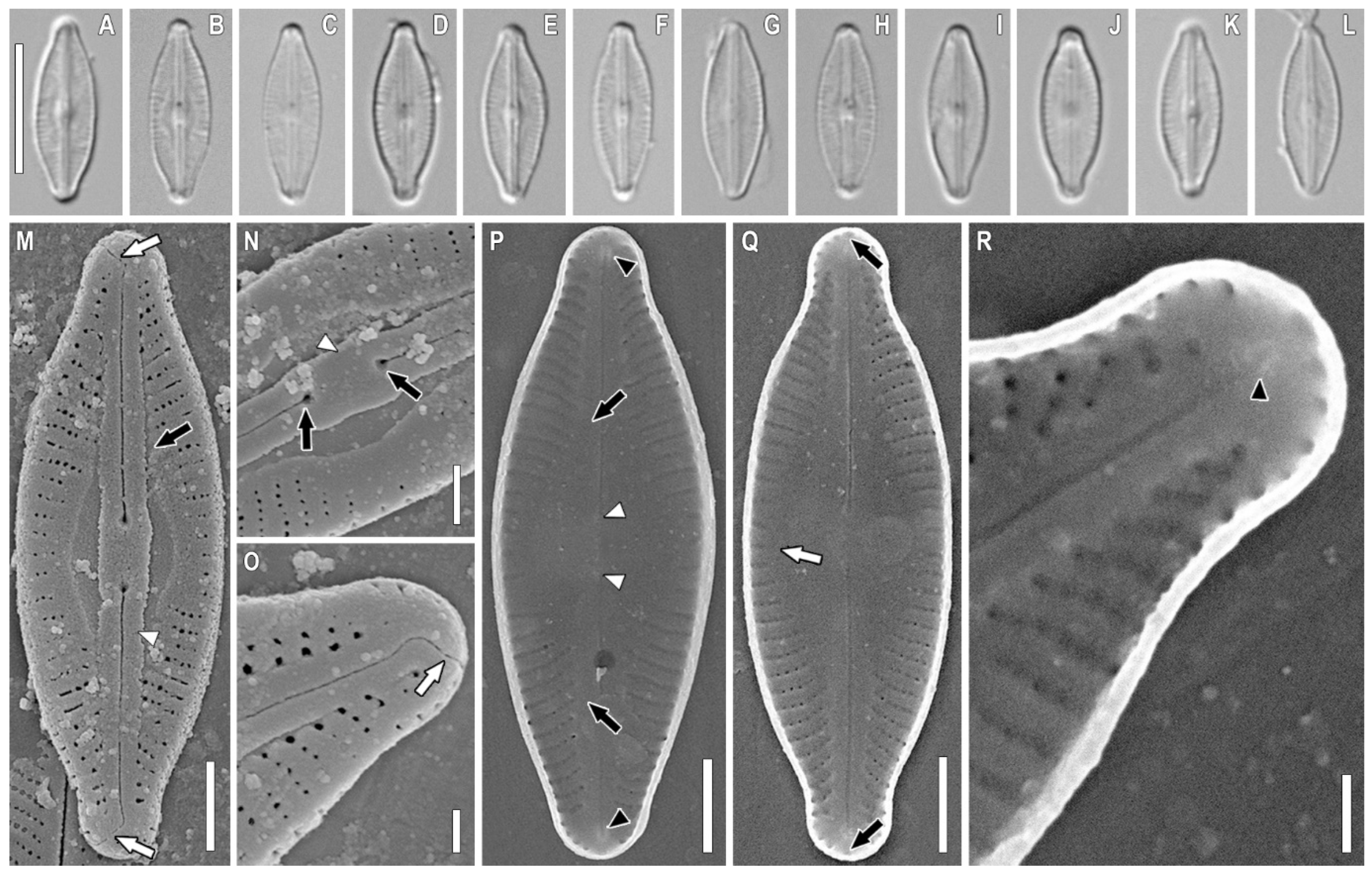
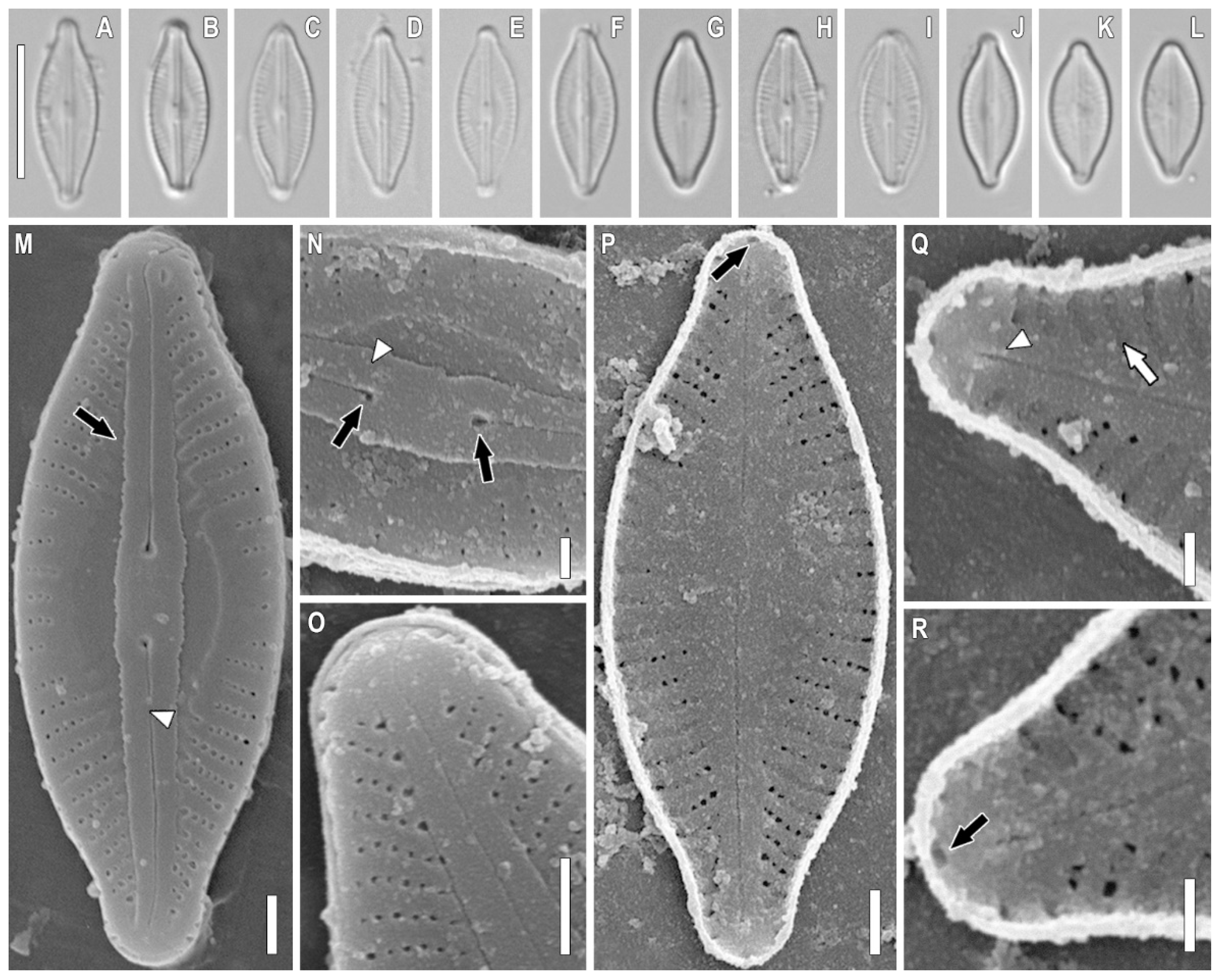
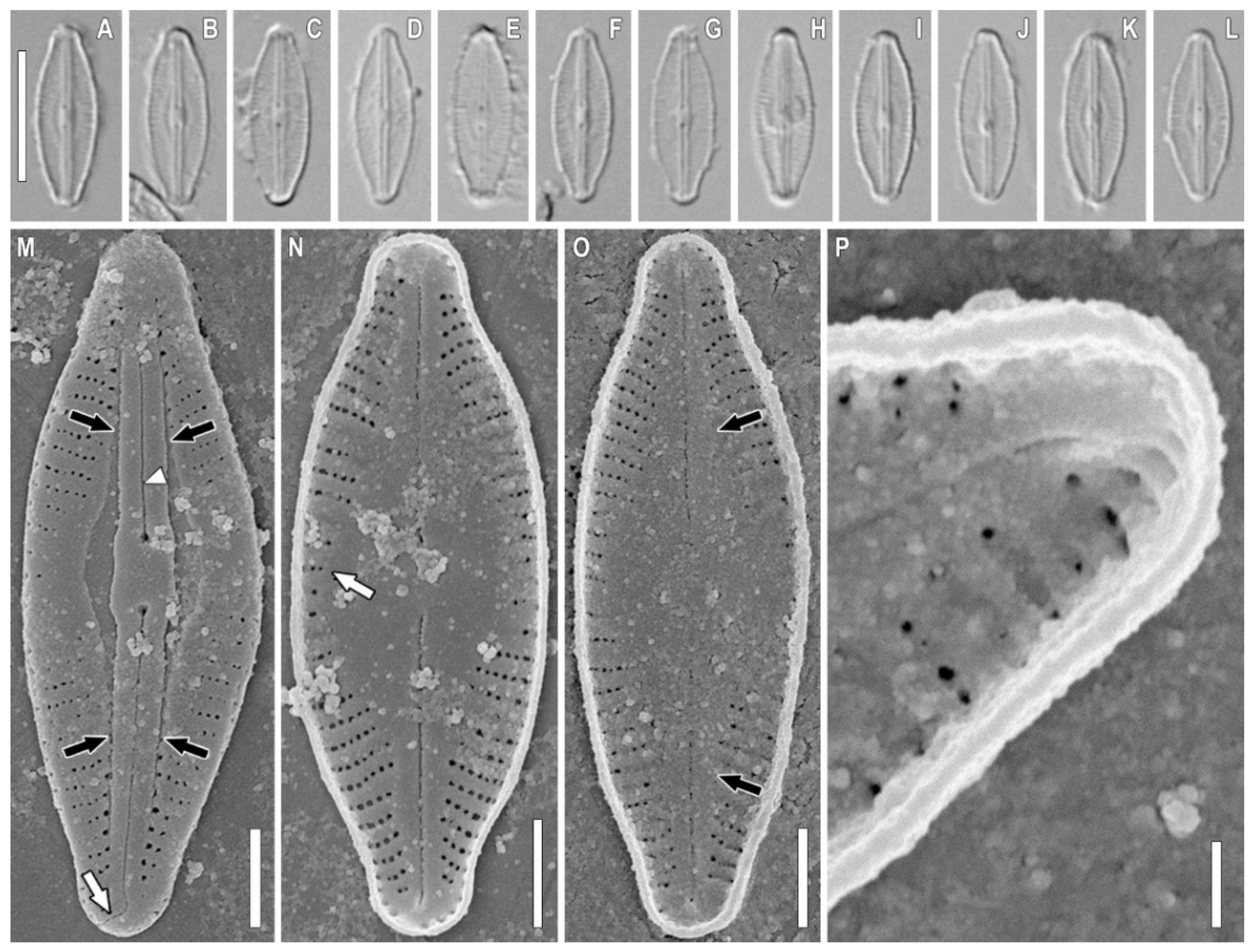
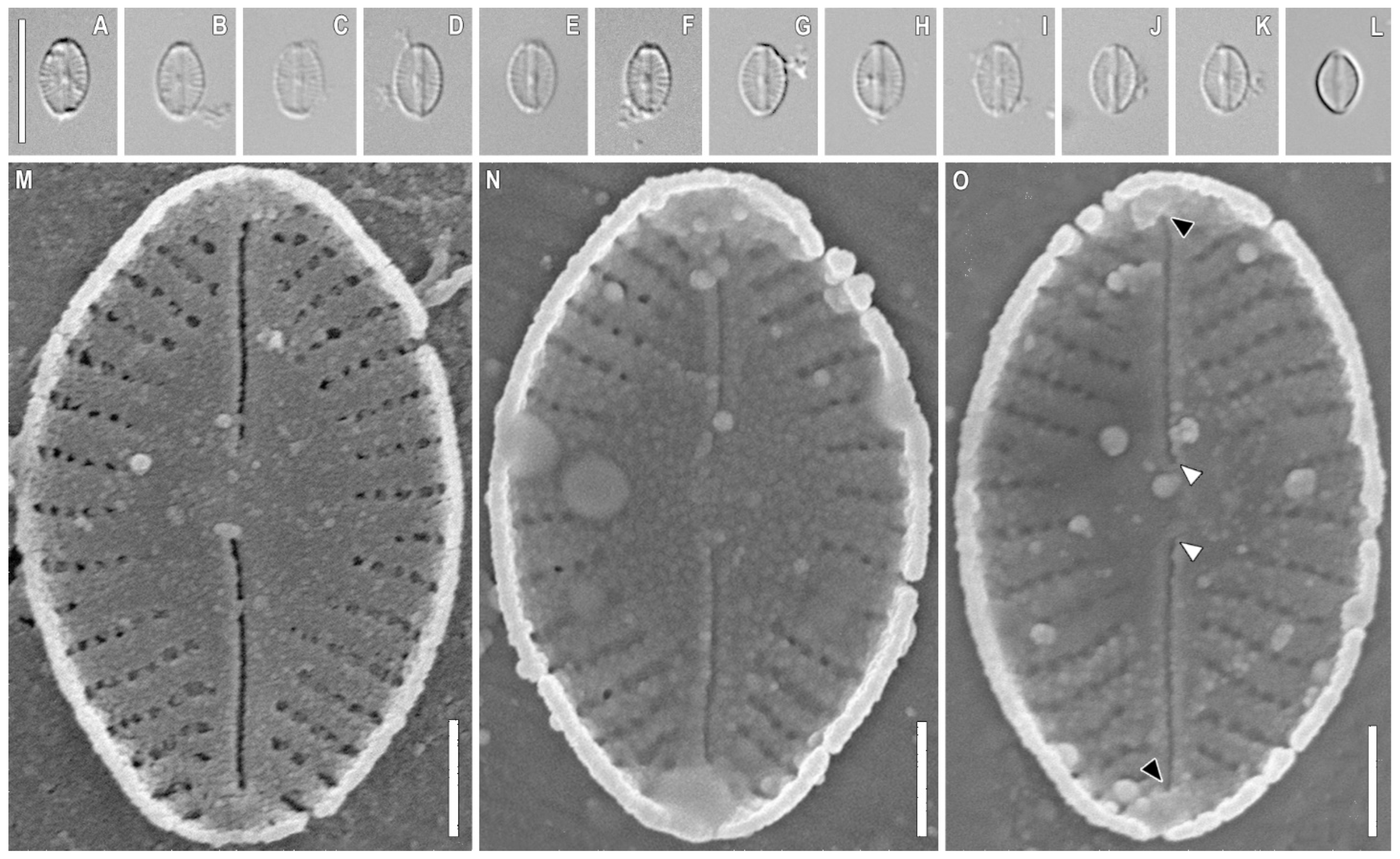
2.1. Taxonomic Analysis
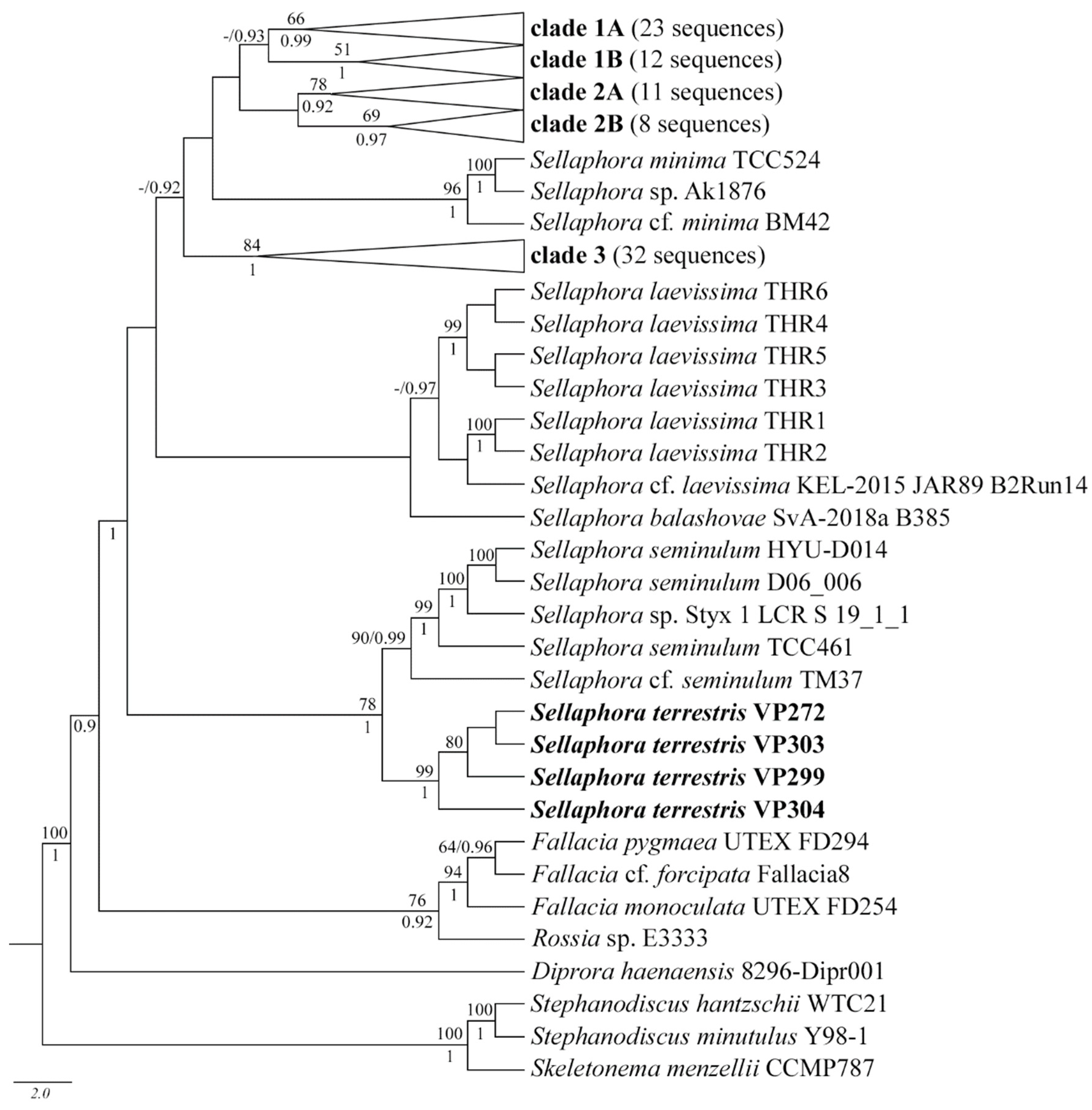
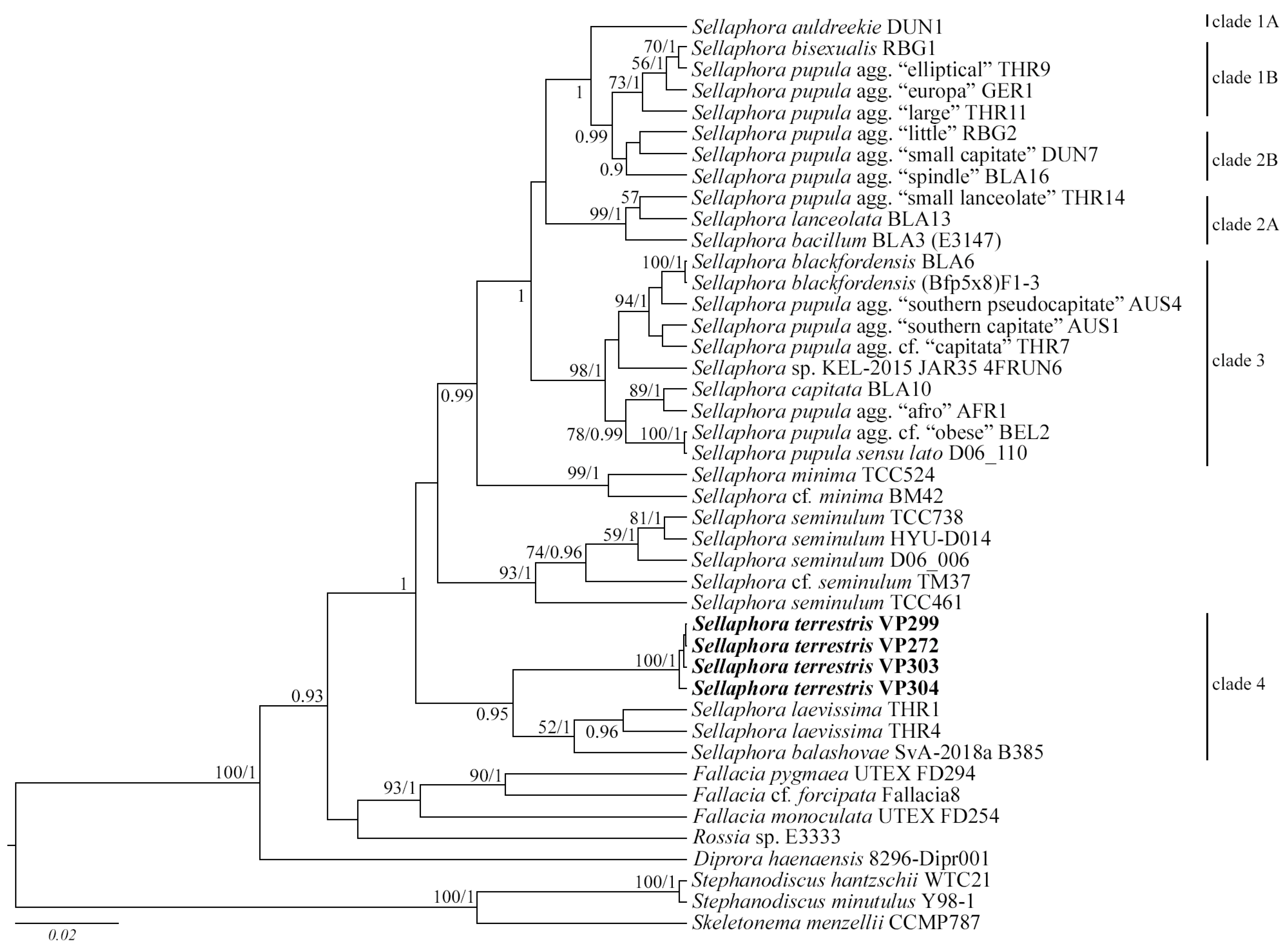
2.2. Molecular Analysis
3. Discussion
3.1. Morphology Discussion
3.2. Molecular Discussion
4. Materials and Methods
4.1. Sample Collection Procedure
4.2. Soil Acidity Measurements
4.3. Culturing
4.4. Preparation of Slides and Microscope Investigation
4.5. Molecular Investigation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mereschkowsky, C. On Sellaphora, a new genus of diatoms. Ann. Mag. Nat. Hist. 1902, 9, 185–195. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2022; Available online: http://www.algaebase.org (accessed on 1 August 2022).
- Kociolek, J.P.; Blanco, S.; Coste, M.; Ector, L.; Liu, Y.; Karthick, B.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.; Potapova, M.; et al. DiatomBase. Available online: https://www.diatombase.org (accessed on 1 August 2022).
- Kulikovskiy, M.S.; Glushchenko, A.M.; Genkal, S.I.; Kuznetsova, I.V. Identification Book of Diatoms from Russia; Filigran: Yaroslavl, Russia, 2016; 804p. [Google Scholar]
- Kulikovskiy, M.S.; Glushchenko, A.M.; Kuznetsova, I.V.; Kociolek, J.P. Description of the new freshwater diatom genus Okhapkinia gen. nov. from Laos (Southeast Asia), with notes on family Sellaphoraceae Mereschkowsky1902. Fottea 2018, 18, 120–129. [Google Scholar] [CrossRef]
- Mann, D.G.; Chepurnov, V.A.; Droop, S.J.M. Sexuality, incompatibility, size variation, and preferential polyandry in natural populations and clones of Sellaphora pupula (Bacillariophyceae). J. Phycol. 1999, 35, 152–170. [Google Scholar] [CrossRef]
- Mann, D.G.; McDonald, S.M.; Bayer, M.M.; Droop, S.J.M.; Chepurnov, V.A.; Loke, R.E.; Ciobanu, A.; du Buf, J.M.H. The Sellaphora pupula species complex (Bacillariophyceae): Morphometric analysis, ultrastructure and mating data provide evidence for five new species. Phycologia 2004, 43, 459–482. [Google Scholar] [CrossRef]
- Mann, D.G.; Thomas, S.J.; Evans, K.M. Revision of the diatom genus Sellaphora: A first account of the larger species in the British Isles. Fottea 2008, 8, 15–78. [Google Scholar] [CrossRef]
- Evans, K.M.; Wortley, A.H.; Simpson, G.E.; Chepurnov, V.A.; Mann, D.G. A molecular systematic approach to explore diversity within the Sellaphora pupula species complex (Bacillariophyta). J. Phycol. 2008, 44, 215–231. [Google Scholar] [CrossRef]
- Mann, D.G.; Stickle, A.S. Cytological characteristics of the Sellaphoraceae. Acta Bot. Croat. 2009, 68, 239–250. [Google Scholar]
- Andreeva, S.; Kociolek, J.P.; Maltsev, Y.; Dorofeyuk, N.; Kezlya, E.; Shkurina, N.; Kuznetsova, I.; Gusev, E.; Kulikovskiy, M.S. Sellaphora balashovae (Bacillariophyta), a new species from Siberian mountain Lake Frolikha (Baikal region), Russia. Phytotaxa 2018, 371, 73–83. [Google Scholar] [CrossRef]
- Mann, D.G.; Poulíčková, A. Homothallism, morphology and phylogenetic position of a new species of Sellaphora (Bacillariophyta), S. pausariae. Plant Ecol. Evol. 2019, 152, 203–218. [Google Scholar] [CrossRef]
- Behnke, A.; Friedl, T.; Chepurnov, V.A.; Mann, D.G. Reproductive compatibility and rDNA sequence analyses in the Sellaphora pupula species complex (Bacillariophyta). J. Phycol. 2004, 40, 193–208. [Google Scholar] [CrossRef]
- Evans, K.M.; Wortley, A.H.; Mann, D.G. An assessment of potential diatom ‘‘barcode’’ genes (cox1, rbcL, 18S and ITS rDNA) and their effectiveness in determining relationships in Sellaphora (Bacillariophyta). Protist 2007, 158, 349–364. [Google Scholar] [CrossRef]
- Evans, K.M.; Chepurnov, V.A.; Sluiman, H.J.; Thomas, S.J.; Spears, B.M.; Mann, D.G. Highly differentiated populations of the freshwater diatom Sellaphora capitata suggest limited dispersal and opportunities for allopatric speciation. Protist 2009, 160, 386–396. [Google Scholar] [CrossRef]
- Mann, D.G. The species concept in diatoms: Evidence for morphologically distinct, sympatric gamodemes in four epipelic species. Plant Syst. Evol. 1989, 164, 215–237. [Google Scholar] [CrossRef]
- Hustedt, F. Die Kieselalgen Deutschlands, Österreichs und der Schweiz unter Berücksichtigung der übrigen Lånder Europas Sowie der Angrenzenden Meeresgebiete. In Österreich und der Schweiz 7 (3:1); von Deutschland, R.K., Ed.; Akademische Verlagsgesellschaft: Leipzig, Germany, 1961; pp. 1–160. [Google Scholar]
- Ross, R. The diatom genus Capartogramma and the identity of Schizostauron. Bull. Br. Mus. Nat. Hist. Bot. 1963, 3, 47–92. [Google Scholar]
- VanLandingham, S.L. Catalogue of the Fossil and Recent Genera and Species of Diatoms and Their Synonyms. Part 5: Navicula; J. Cramer: Vaduz, Liechtenstein, 1975; pp. 386–963. [Google Scholar]
- Vanormelingen, P.; Evans, K.M.; Chepurnov, V.A.; Vyverman, W.; Mann, D.G. Molecular species discovery in the diatom Sellaphora and its congruence with mating trials. Fottea 2013, 13, 133–148. [Google Scholar] [CrossRef]
- Poulíčková, A.; Sato, S.; Evans, K.M.; Chepurnov, V.A.; Mann, D.G. Repeated evolution of uniparental reproduction in Sellaphora (Bacillariophyceae). Eur. J. Phycol. 2015, 50, 62–79. [Google Scholar] [CrossRef]
- Hamsher, S.E.; Evans, K.; Mann, D.G.; Poulíčková, A.; Saunders, G.W. Barcoding Diatoms: Exploring Alternatives to COI-5P. Protist 2011, 162, 405–422. [Google Scholar] [CrossRef]
- Hustedt, F. Bacillariophyta (Diatomeae). In Die Süsswasser-Flora Mitteleuropas. Zweite Auflage. Heft 10; Pascher, A., Ed.; Gustav Fischer: Jena, Germany, 1930; 466p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae. In Süβwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer: Stuttgart, Germany, 1986; Volume 2, 876p. [Google Scholar]
- Mann, D.G. Observations on copulation in Navicula pupula and Amphora ovalis in relation to the nature of diatom species. Ann. Bot. 1984, 54, 429–438. [Google Scholar] [CrossRef]
- Mann, D.G.; Evans, K.M.; Chepurnov, V.A.; Nagai, S. Morphology and formal description of Sellaphora bisexualis sp. nov. (Bacillariophyta). Fottea 2009, 9, 199–209. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Barragán, C.; Ector, L. Sellaphora lundii nom. et stat. nov. (Bacillariophyta), a forgotten European terrestrial species. Not. Algarum 2017, 38, 1–3. [Google Scholar]
- Foets, J.; Wetzel, C.E. Sellaphora harderi (Hustedt) comb. nov. (Bacillariophyta, Sellaphoraceae), an overlooked terrestrial diatom. Not. Algarum 2018, 52, 1–5. [Google Scholar]
- Kuehnle, A.; Schurr, R.; Perez, M. Some Microalgae from the Hawaiian Islands with a Focus on Industrial Applications. Curr. Biotechnol. 2015, 4, 499–513. [Google Scholar] [CrossRef]
- Malavasi, V.; Škaloud, P.; Rindi, F.; Tempesta, S.; Paoletti, M.; Pasqualetti, M. DNA-based taxonomy in ecologically versatile microalgae: A re-evaluation of the species concept within the coccoid green algal genus Coccomyxa (Trebouxiophyceae, Chlorophyta). PLoS ONE 2016, 11, e0151137. [Google Scholar] [CrossRef]
- Kezlya, E.M.; Glushchenko, A.M.; Maltsev, Y.I.; Maltseva, S.Y.; Gusev, E.S.; Kulikovskiy, M.S. Composition and structure algal Community of the Cát Tiên National Park, southern Vietnam (first data). Issues Mod. Algol. 2021, 2, 78–85. [Google Scholar] [CrossRef]
- Kezlya, E.M.; Glushchenko, A.M.; Kociolek, J.P.; Maltsev, Y.I.; Martynenko, N.A.; Genkal, S.I.; Kulikovskiy, M.S. Mayamaea vietnamica sp. nov.: A new, terrestrial diatom (Bacillariophyceae) species from Vietnam. Algae 2020, 35, 325–335. [Google Scholar] [CrossRef]
- Kezlya, E.M.; Glushchenko, A.M.; Maltsev, Y.I.; Gusev, E.S.; Genkal, S.I.; Kuznetsov, A.N.; Kociolek, J.P.; Kulikovskiy, M.S. Placoneis cattiensis sp. nov.—A new, diatom (Bacillariophyceae: Cymbellales) soil species from Cát Tiên National Park (Vietnam). Phytotaxa 2020, 460, 237–248. [Google Scholar] [CrossRef]
- Falasco, E.; Blanco, S.; Bona, F.; Goma, J.; Hlubikova, D.; Novais, M.H.; Hoffmann, L.; Ector, L. Taxonomy, morphology and distribution of the Sellaphora stroemii complex (Bacillariophyceae). Fottea 2009, 9, 243–256. [Google Scholar] [CrossRef]
- Tuji, A. Freshwater diatom flora in the bottom sediments of Lake Biwa (South Basin): Navicula sensu lato. Bull. Natl. Sci. Mus. Tokyo Ser. B Bot. 2003, 29, 65–82. [Google Scholar]
- Hustedt, F. Systematische und ökologische Untersuchungen über die Diatomeen-Flora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda-Expedition “Tropische Binnengewässer, Band VII”. Arch. Für Hydrobiol. Suppl. 1937, 15, 187–295. [Google Scholar]
- Wetzel, C.E.; Ector, L.; Van de Vijver, B.; Compère, P.; Mann, D.G. Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea 2015, 15, 203–234. [Google Scholar] [CrossRef]
- Cantonati, M.; Kelly, M.G.; Lange-Bertalot, H. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Königstein, Germany, 2017; 942p. [Google Scholar]
- Heudre, D.; Wetzel, C.E.; Moreau, L.; Ector, L. Sellaphora davoutiana sp. nov.: A new freshwater diatom species (Sellaphoraceae, Bacillariophyta) in lakes of Northeastern France. Phytotaxa 2018, 346, 269–279. [Google Scholar] [CrossRef]
- Lund, J.W.G. Observations on soil algae I. The ecology, size and taxonomy of British soil diatoms. Part 2. New Phytol. 1946, 45, 56–110. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Ector, L.; Haan, M.; Zidarova, R. The genus Microcostatus in the Antarctic region. Diatom Res. 2010, 25, 417–429. [Google Scholar] [CrossRef]
- Zidarova, R.; Kopalová, K.; Van de Vijver, B. Diatoms from the Antarctic Region: Maritime Antarctica. Iconogr. Diatomol. 2016, 24, 1–504. [Google Scholar]
- Stanek-Tarkowska, J.; Czyz, E.A.; Rybak, M. Description of a new diatom species—Microcostatus dexteri sp. nov.—From terrestrial habitats in southern Poland. Phytotaxa 2021, 509, 241–247. [Google Scholar] [CrossRef]
- Johansen, J.R.; Sray, J.C. Microcostatus gen. nov., a new aerophilic diatom genus based on Navicula krasskei Hustedt. Diatom Res. 1998, 13, 93–101. [Google Scholar] [CrossRef]
- Liu, Y.; Kociolek, J.P.; Lu, X.; Fan, Y. A new Sellaphora Mereschkowsky species (Bacillariophyceae) from Hainan Island, China, with comments on the current state of the taxonomy and morphology of the genus. Diatom Res. 2020, 35, 85–98. [Google Scholar] [CrossRef]
- Tuji, A.; Suzuki, S.; Yamagishi, T.; Yamaguchi, H. Alternative Diatom Strains for Growth Inhibition Tests (OECD Test Guideline 201). Bull. Natl. Mus. Nat. Sci. Ser. B Bot. 2021, 47, 183–192. [Google Scholar] [CrossRef]
- Blanc, L.; Maury-Lechon, G.; Pascal, J.-P. Structure, floristic composition and natural regeneration in the forests of Cat Tien National Park, Vietnam: An analysis of the successional trends. J. Biogeogr. 2001, 27, 141–157. [Google Scholar] [CrossRef]
- Vadjunina, A.F.; Korchagina, Z.A. Methods of Studying the Physical Properties of Soils; Agropromisdat: Moscow, Russia, 1986; 416p. (In Russian) [Google Scholar]
- Arinushkina, E.W. Handbook for Chemical Soil Analysis; Publishing Moscow State University: Moscow, Russia, 1970; 488p. (In Russian) [Google Scholar]
- McFadden, G.I.; Melkonian, M. Use of Hepes buffer for microalgal culture media and fixation for electron microscopy. Phycologia 1986, 25, 551–557. [Google Scholar] [CrossRef]
- Zimmermann, J.; Jahn, R.; Gemeinholzer, B. Barcoding diatoms: Evaluation of the V4 subregion on the 18S rRNA gene, including new primers and protocols. Org. Divers. Evol. 2011, 11, 173–192. [Google Scholar] [CrossRef]
- Ruck, E.C.; Theriot, E.C. Origin and evolution of the canal raphe system in diatoms. Protist 2011, 162, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Alverson, A.J.; Jansen, R.K.; Theriot, E.C. Bridging the Rubicon: Phylogenetic analysis reveals repeated colonization’s of marine and fresh waters by thalassiosiroid diatoms. Mol. Phylogenet. Evol. 2007, 45, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Met. 2012, 9, 772. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild Population | Culture | ||||||
|---|---|---|---|---|---|---|---|
| Sample Kt 33 | Sample Kt 59 | Sample Kt 60 | Strain VP 272 | Strain VP 299 | Strain VP 303 | Strain VP 304 | |
| Outline | linear-lanceolate, linear-elliptical to elliptical | linear-lanceolate, linear-elliptical to elliptical | linear-elliptical to elliptical | linear-elliptical to elliptical | elliptical | linear-elliptical | linear-elliptical |
| Ends | short rostrate to broadly rounded | short rostrate to broadly rounded | short rostrate to broadly rounded | rostrate | short rostrate to broadly rounded | rostrate | rostrate |
| Axial area | narrow | narrow | narrow | narrow | narrow | narrow | narrow |
| Central area | small, elliptical | small, elliptical | small, elliptical | small, elliptical | small, elliptical | small, elliptical | small, elliptical |
| Valve length (μm) | 9.1–17.1 (11.7 ± 2.4; n = 22) | 8.2–16.9 (12.3 ± 2.3; n = 22) | 6.9–13.4 (9.5 ± 1.4; n = 22) | 10.4–13.9 (11.7 ± 0.8; n = 22) | 5.2–6.3 (5.5 ± 0.2; n = 21) | 13.2–14.0 (13.5 ± 0.2; n = 23) | 13.4–14.3 (13.8 ± 0.2; n = 24) |
| Valve width (μm) | 4.5–6.1 (4.9 ± 0.4; n = 22) | 4.2–5.0 (4.5 ± 0.2; n = 22) | 4.3–5.0 (4.4 ± 0.2; n = 22) | 4.5–4.9 (4.7 ± 0.1; n = 22) | 3.7–4.0 (3.8 ± 0.05; n = 21) | 4.7–4.9 (4.8 ± 0.03; n = 23) | 4.6–5.1 (4.8 ± 0.1; n = 24) |
| Striae, in 10 μm | 29–32 (28.6 ± 1.3; n = 22) | 30–31 (30.1 ± 0.3; n = 22) | 29–31 (29.9 ± 0.4; n = 22) | 29–30 (29.9 ± 0.2; n = 22) | 30–34 (32 ± 0.7; n = 21) | 28–32 (30 ± 0.6; n = 23) | 30–35 (32.1 ± 0.9; n = 24) |
| Areolae | - | - | - | - | ca. 65 | ca. 60 | ca. 60 |
| Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Sellaphora terrestris VP304 | – | |||||||||||||||||||
| 2 | Microcostatus sp. MMDL 54702 | 95.4 | – | ||||||||||||||||||
| 3 | Microcostatus sp. MMDL 54701 | 91.8 | 92.9 | – | |||||||||||||||||
| 4 | Sellaphora auldreekie DUN1 clade_1A | 96.4 | 94.4 | 90.8 | – | ||||||||||||||||
| 5 | Sellaphora pupula agg. “large” THR11 clade 1B | 95.7 | 95.2 | 90.8 | 97.7 | – | |||||||||||||||
| 6 | Sellaphora bisexualis RBG1 clade 1B | 95.9 | 95.4 | 91.1 | 98.0 | 99.8 | – | ||||||||||||||
| 7 | Sellaphora lanceolata BLA13 clade 2A | 96.4 | 94.4 | 90.0 | 97.4 | 96.7 | 96.9 | – | |||||||||||||
| 8 | Sellaphora bacillum BLA3 clade 2A | 96.7 | 95.2 | 90.8 | 97.7 | 97.0 | 97.2 | 99.2 | – | ||||||||||||
| 9 | Sellaphora pupula agg. “little” RBG clade 2B | 95.4 | 95.4 | 91.1 | 97.5 | 99.5 | 99.8 | 96.4 | 96.7 | – | |||||||||||
| 10 | Sellaphora pupula agg. “small capitate” DUN7 clade 2B | 95.9 | 95.4 | 91.1 | 98.0 | 99.8 | 99.7 | 97.2 | 97.5 | 99.5 | – | ||||||||||
| 11 | Sellaphora blackfordensis BLA6 clade 3 | 95.9 | 95.1 | 90.8 | 97.9 | 97.2 | 97.4 | 97.2 | 97.7 | 96.9 | 97.4 | – | |||||||||
| 12 | Sellaphora pupula agg. cf. obesa BEL2 clade 3 | 95.4 | 94.9 | 90.5 | 97.2 | 96.9 | 97.2 | 96.4 | 96.9 | 96.7 | 97.2 | 99.2 | – | ||||||||
| 13 | Sellaphora balashovae SvA-2018a B385 | 96.7 | 95.7 | 91.3 | 96.9 | 96.4 | 96.7 | 96.7 | 97.2 | 96.2 | 96.7 | 96.9 | 96.2 | – | |||||||
| 14 | Sellaphora sp. KEL-2015 JAR35 4FRun6 | 95.9 | 94.6 | 90.3 | 97.4 | 96.9 | 97.2 | 96.7 | 97.2 | 96.7 | 97.2 | 98.5 | 97.7 | 96.9 | – | ||||||
| 15 | Sellaphora minima TCC524 | 96.2 | 93.9 | 90.8 | 95.9 | 95.2 | 95.4 | 95.4 | 95.7 | 95.4 | 95.4 | 94.9 | 94.1 | 95.7 | 94.9 | – | |||||
| 16 | Sellaphora laevissima THR4 | 97.2 | 96.7 | 91.8 | 96.9 | 97.2 | 97.4 | 97.2 | 97.7 | 96.9 | 97.4 | 98.0 | 97.4 | 98.5 | 98.0 | 95.9 | – | ||||
| 17 | Sellaphora laevissima THR1 | 96.4 | 95.9 | 91.1 | 96.2 | 96.4 | 96.7 | 96.4 | 96.9 | 96.2 | 96.7 | 97.2 | 96.7 | 97.7 | 97.2 | 94.9 | 99.2 | – | |||
| 18 | Sellaphora seminulum HYU-D014 | 94.9 | 92.6 | 89.0 | 95.2 | 94.4 | 94.6 | 95.9 | 95.9 | 94.2 | 94.9 | 94.9 | 94.2 | 94.9 | 93.9 | 93.7 | 94.9 | 94.7 | – | ||
| 19 | Sellaphora seminulum TCC461 | 94.1 | 93.4 | 89.0 | 93.6 | 93.9 | 94.1 | 93.6 | 93.6 | 93.6 | 94.4 | 94.1 | 93.9 | 94.1 | 93.6 | 93.9 | 95.4 | 94.6 | 95.7 | – | |
| 20 | Sellaphora cf. seminulum TM37 | 95.9 | 94.4 | 90.8 | 96.7 | 96.2 | 96.4 | 96.7 | 97.0 | 96.4 | 96.4 | 96.5 | 95.7 | 96.9 | 95.4 | 95.4 | 96.4 | 95.7 | 96.7 | 94.7 | – |
| Outline | Ends | Axial Area | Central Area | Valve Length (μm) | Valve Width (μm) | Striae in 10 μm | Areolae in 10 μm | Distribution | References | |
|---|---|---|---|---|---|---|---|---|---|---|
| Sellaphora terrestris sp. nov. | linear-elliptical to elliptical with weakly convex margins | short rostrate to broadly rounded | narrow, weakly extended to the central area | small, longitudinally lanceolate | 6.9–17.1 | 3.7–6.1 | 28–35 | 60–65 | Vietnam, Cát Tiên National Park | This study |
| S. laevissima | linear, usually with weakly convex margins in the centre | either very weakly protracted, subcapitate or not protracted, always broadly rounded | narrow, not or very little broadened towards the centre | distinct, more or less transversely rectangular, extends over 50–60% of the valve width, rarely more | 22–50 | 7–11 | 15–21 | 30–35 | Holarctic | [38] |
| S. balashovae | elliptic | subcapitate | narrow, weakly extended to the central area | bow-tie-shaped, bordered by alternating short and long striae | 11–14 | 4 | 30 | 60 | Russia, Lake Frolikha | [11] |
| S. seminulum | linear–lanceolate to broadly lanceolate with clearly convex margins (inflated in the middle) | protracted, broadly rounded, slightly rostrate | linear–lanceolate, with an irregular border, and is central or apparently very slightly displaced towards the secondary side | bordered by three shorter striae and is usually butterfly–shaped, but the size and shape vary | 5.0–16.5 | 3.0–4.5 | 18–20 | >30 | Holarctic | [37,38] |
| S. davoutiana | elliptic-lanceolate | broadly protracted, subcapitate and rounded | narrow, linear and slightly widened toward central area | small and elliptical to rectangular with 2 or 3 slightly shortened striae of varying length on either side | 28–35 | 4.0–4.9 | 26–30 | 60–70 | France. Grand Est region. La Bresse: Lispach Lake | [39] |
| S. lundii | elliptical, are centrally inflated | rostrate to weakly capitate | narrow, linear | clearly marked, rectangular to oblong | 9–13 | 4–5 | 23–28 | 55–60 * | England, UK, Selly Oak, Birmingham. Allotment; Belgium, Luxembourg | [27,40] |
| Microcostatus naumanii | elliptic-lanceolate to lanceolate with clearly convex margins | in smaller specimens, apices tend to be more rostrate, whereas larger valves have protracted, capitate ends | very narrow near the valve apices but forming two longitudinal depressions, abruptly and elliptically enlarged towards the central area | almost elliptical | 9.5–15.5 | 4–5 | 24–26 | c.a. 60 * | Sweden, Macedonia, Norway, Finland, Victoria Island, Scotland, Patagonia (Argentina), South Shetland Islands (Deception Island, King George Island and Livingston Island), Antarctic Continent | [41,42] |
| Microcostatus dexterii | lanceolate to elliptical-lanceolate | rounded | narrowing near the valve apices, forming two longitudinal depressions, enlarged in the middle and creating an central area | almost lanceolate | 6.6–16.4 | 2.3–5.5 | 20–24 | 45–50 * | Poland. Podkarpacie Province, Rzeszów, soils | [43] |
| Strains | Slide No | Sample No | Sample Locality | Collection of Date | Collection Site | pH | Total Humidity (%) | GenBank Accession Number, SSU rDNA, Partial | GenBank Accession Number, rbcL, Partial |
|---|---|---|---|---|---|---|---|---|---|
| Sellaphora terrestris sp. nov. VP 272 | 07030 | Kt33 | Southeast Vietnam, Cat Tien National Park | 25 June 2012 | 11°25.019′ N 107°25.545′ E Forest soil surface | 6.0 | 48.97 | ON332054 | ON350763 |
| Sellaphora terrestris sp. nov. VP 299 | 07057 | Kt60 | Southeast Vietnam, Cat Tien National Park | 25 June 2019 | 11°24.247′ N 107°23.661′ E Agricultural field soil surface | 4.58 | 27.9 | ON332055 | ON350764 |
| Sellaphora terrestris sp. nov. VP 303 | 07061 | Kt59 | Southeast Vietnam, Cat Tien National Park | 25 June 2019 | 11°24.241′ N 107°22.468′ E Agricultural field soil surface | 5.22 | 36 | ON332056 | ON350765 |
| Sellaphora terrestris sp. nov. VP 304 | 07062a | Kt59 | Southeast Vietnam, Cat Tien National Park | 25 June 2019 | 11°24.241′ N 107°22.468′ E Agricultural field soil surface | 5.22 | 36 | ON332057 | ON350766 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glushchenko, A.; Kezlya, E.; Maltsev, Y.; Genkal, S.; Kociolek, J.P.; Kulikovskiy, M. Description of the Soil Diatom Sellaphora terrestris sp. nov. (Bacillariophyceae, Sellaphoraceae) from Vietnam, with Remarks on the Phylogeny and Taxonomy of Sellaphora and Systematic Position of Microcostatus. Plants 2022, 11, 2148. https://doi.org/10.3390/plants11162148
Glushchenko A, Kezlya E, Maltsev Y, Genkal S, Kociolek JP, Kulikovskiy M. Description of the Soil Diatom Sellaphora terrestris sp. nov. (Bacillariophyceae, Sellaphoraceae) from Vietnam, with Remarks on the Phylogeny and Taxonomy of Sellaphora and Systematic Position of Microcostatus. Plants. 2022; 11(16):2148. https://doi.org/10.3390/plants11162148
Chicago/Turabian StyleGlushchenko, Anton, Elena Kezlya, Yevhen Maltsev, Sergei Genkal, John Patrick Kociolek, and Maxim Kulikovskiy. 2022. "Description of the Soil Diatom Sellaphora terrestris sp. nov. (Bacillariophyceae, Sellaphoraceae) from Vietnam, with Remarks on the Phylogeny and Taxonomy of Sellaphora and Systematic Position of Microcostatus" Plants 11, no. 16: 2148. https://doi.org/10.3390/plants11162148